Introduction
The prevalence of obesity in children and adolescents continues to increase worldwide. The prevalence of obesity in infants and preschool children in South Korea has increased about 2 times in 10 years1). Recently, several studies have reported that obesity affects the incidence of inflammatory diseases2). Furthermore, studies have suggested that obesity induces reduction of the moisturizing and barrier functions of the stratum corneum (SC)3).
Atopic dermatitis (AD) is the most common inflammatory disease of the skin based on the skin barrier dysfunction characterized by pruritus, dryness, and erythematous eczema4). Recently the hypothesis that the pathogenesis of AD is caused by the destruction of the skin barrier system is emerging5). The defects of skin barrier induced a various abnormalities in lipid composition and the extent of the permeability barrier abnormality in AD6). Thus the abnormalities of the skin barrier are not simply epiphenomenon of AD, but in preference the driver for inflammation of AD7). Damages to skin barrier function have highly interrelation with inflammatory skin disease induction. Recently, Interleukin (IL)-31 that is a cytokine associated with Th2 differentiation in AD, has been shown to reduce the expression of filaggrin, a major constituent of the skin barrier8). In addition, reported that the mutated or reduced filaggrin can cause inflammation by damaging the skin barrier9).
Within traditional Korean medicine, AD is regarded as related to a fetal heat or fetal poisoning, which are caused by unhealthy dietary habits such as excessive fat intake during pregnancy10). Hataedock (HTD) is traditional treatments of Korean medicine in which herbal extracts are orally administered to neonates to prevent inflammatory diseases caused by mother’s dietary habits during pregnancy. Coptidis Rhizoma, Glycyrrhizae Radix (CRGR) are a representative herbal medicine of HTD treatments. Our previous study suggested that HTD treatment maintains and fortifies the lipid barrier11). Based on that study, we sought to study the effect of HTD treatment on the maintenance of lipid barrier in obese conditions that could promote an inflammatory response.
In this study, CRGR was administered orally to 3-week-old NC/Nga mice as the HTD treatments and pediatric obese condition was induced by high fat diet. To investigate the lipid barrier maintenance effect of HTD treatment, Dermatophagoides farinae extract (DfE) was applied to NC/Nga mice to induce AD-like skin lesions. In a series of pathway related to provoking AD, HTD treatment is anticipated to assist in maintenance of the lipid barrier. Thus, we investigated the maintenance effects of HTD treatments on the lipid barrier disrupted by DfE in AD-induced obese NC/Nga mice.
Materials and Methods
1. Preparation of extracts
In this study, extracts of Coptidis Rhizoma (Coptis Japonica) and Glycyrrhizae Radix (Glycyrrhiza uralensis) were used for HTD treatment. The CRGR extract was prepared as follows: 1) Coptidis Rhizoma (100 g) and Glycyrrhizae Radix (100 g) were decocted in 1 L of distilled water for 3 hours and then filtered; 2) The decoction was concentrated to 50 mL by a rotatory vacuum evaporator, and the filtrate was freeze-dried. As a result, we obtained 31 g of extract (yield: 15.5%).
2. Experimental animals and induction of AD
Male 3-week-old NC/Nga mice, which were 14.3±0.3 g in weight, were obtained from Central Lab. Animal Inc. (Seoul, Republic of Korea). For diet-induced obesity, diet composed of 60% fat, 20% carbohydrates, and 20% protein was administered. All mice were allowed to eat freely throughout the experiment.
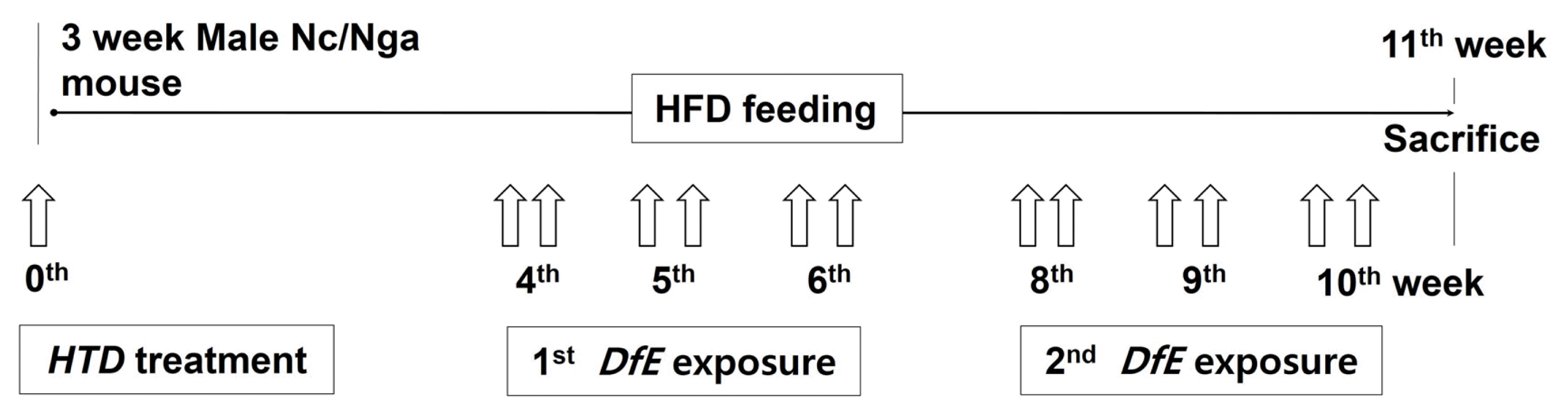
Each 10 mice were allocated to three groups (total 30 mice) as follows: the high fat diet group (Ctrl group), high fat diet and AD-induced group (AE group), and high fat diet and AD-induced group after HTD treatment with CRGR extract group (CGT group). In CGT group, with 3-week-old mice, HTD treatments that CRGR extracts (20 mg/kg each) are orally administered to each group on the 1st, 2nd, and 3rd days was performed. For AD-like skin lesion, we skinned the backs of mice and swabbed the skinned areas 20 times with 1 mL of 5% sodium dodecyl sulfate (SDS; Sigma-Aldrich, St. Louis, MO, USA) using a cotton swab to remove the lipid lamella of the SC. Then, 100 mg of DfE (Biostir Inc., Kobe, Japan) was applied to the mice two times per week. The first exposure was conducted on the 4th, 5th, and 6th weeks. After 1 week, the second exposure was conducted on the 8th, 9th, and 10th weeks. On the 11th week, mice were sacrificed with sodium pentobarbital. The overall procedure is depicted in Fig 1.
All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of Pusan National University (IACUC number: PNU-2015-0924). We also followed the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals throughout this study.
3. Tissue sample preparation and histochemistry
After sacrificing the mice, we obtained dorsal skins and then fixed them in 10% neutral-buffered formalin (NBF) at room temperature for 24 hours. The fixed tissues were embedded in paraffin for serial sectioning with 5-μm thickness. We conducted H&E (Hematoxylin and Eosin) staining to investigate histological changes including epithelial hyperplasia, capillary distribution, and collagen fiber distribution of the tissue samples.
4. Immunohistochemistry
Skin tissue slices were soaked in 20 μg/mL of proteinase K solution for 5 minutes for proteolysis. The proteolyzed slices were incubated in 10% normal goat serum for 4 hours, which was used as blocking serum. Next, the slices were treated with primary antibodies (All antibodies used in the experiment were purchased at Santa Cruz Biotechnology, Dallas, TX, USA), including goat anti-Lass2 (1:100), goat anti-filaggrin (1:100), goat anti-involucrin (1:50), goat anti-loricrin (1:50), goat anti-ASM (1:50), goat anti-liver X receptor (LXR; 1:200), goat anti-protein kinase C (1:100), goat anti-IL-4 (1:100), and goat anti-STAT6 (1:100) for 72 hours in a 4 °C humidified chamber.
Then, the slices reacted with the primary antibodies were treated with biotinylated rabbit anti-goat IgG (1:100) secondary antibody for 24 hours at room temperature. After reaction with the secondary antibody, the slices were treated with an avidin-biotin complex kit (Vector Laboratories, Burlingame, CA, USA) for 1 hour at room temperature. As a final step, the slices were developed with 0.05M tris-HCl buffer solution (pH 7.4) consisting of 0.05% 3,3′-diaminobenzidine (DAB) and 0.01% HCl and then counterstained with hematoxylin.
5. Image analysis and statistical analysis
To obtain numerical data from immunohistochemistry, image analysis was performed by Image Pro Plus (Media cybernetics, Rockville, MD, USA). In the image analysis of 400x-magnified exposure photography, positively reacted particle as pixel cells (intensity range: 80–100) were counted from 10 randomly selected fields from each group. Total pixel cells were 20,000,000 depending on the results of immunohistochemistry such as non-specific structure and artificiality. Data were presented as the mean ± standard error (mean ± SE). One-way ANOVA and Post-Hoc test were used to analyze statistical significance of the differences with a significance level of P<0.05. SPSS 23 software (IBM Corp, Armonk, NY, USA) was used for statistical analysis.
Results
1. Alleviating effect on symptoms of AD
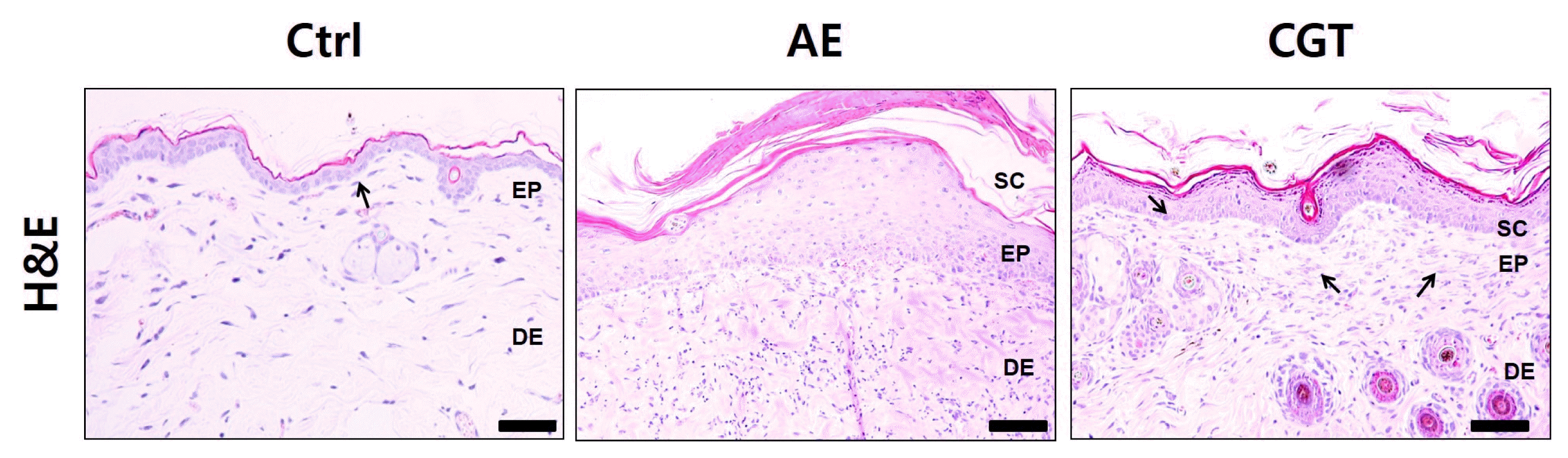
H&E staining was used to observe changes caused by edema including collagen fiber distribution and epithelial hyperplasia. The AE group exhibited pathological change such as reduction in collagen fiber and an increase in epithelial hyperplasia. These results represent the typical histological appearance of inflammatory skin damage induced through persistent application of DfE. In contrast, the CGT group showed less pathological changes than the AE group in most areas. The CGT group exhibited low epidermal hyperplasia and maintenance of collagen fiber density (Fig 2).
2. Maintenance of lipid barrier
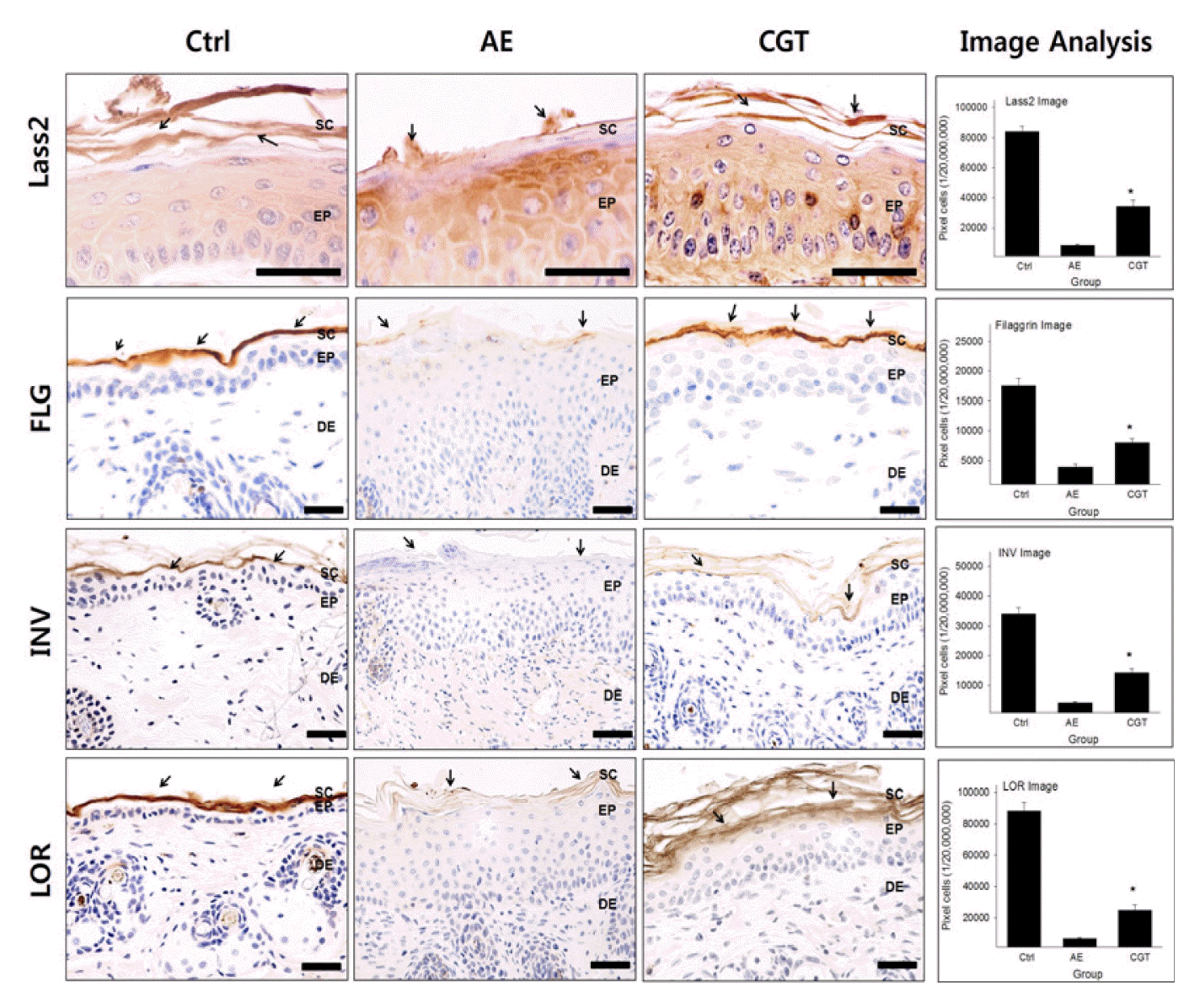
To investigate the protective effect of HTD treatment on the epidermal barrier, Lass2-, filaggrin-, involucrin-, and, loricrin-positive reactions were measured.
Lass2-positive reactions were used to detect the ceramide synthesis of the epidermal lipid layer. The Lass2 level of the AE group was 8,317±221/20,000,000 pixel which was decreased in the SC by 90% compared with that of the Ctrl group. Compared with the AE group, Lass2 levels of the CGT group was increased significantly. The Lass2 level of the CGT group was 34,041±1320/20,000,000 pixel which was increased by 309% (P<0.05) compared with that of the AE group (Fig 3).
Filaggrin-positive reactions were mainly found in keratohyalin granule of SC. The filaggrin level of the AE group was 5,511±390/20,000,000 pixel which was decreased by 49% compared with that of the Ctrl group. However, both HTD treatments recovered filaggrin levels in the CGT group. The levels of filaggrin in the CGT group was respectively 8,840±218/20,000,000 pixel which were increased by 60% (P<0.05) compared with that of the AE group (Fig 3).
Involucrin-positive reactions were observed in cornified layer of SC. The involucrin level of the AE group was 4,417±93/20,000,000 pixel which was decreased by 87% compared with that of the Ctrl group. In contrast, HTD treatments improved involucrin levels in the CGT group. The levels of involucrin in the CGT group was 14,342±419/20,000,000 pixel which were significantly increased by 225% (P<0.05) compared with that of the AE group (Fig 3).
The results for loricrin-positive reaction were similar to the results for involucrin. Compared with the Ctrl group, the loricrin level of the AE group was 6,501±227/20,000,000 pixel which was decreased by 93%. After both HTD treatments, the loricrin levels increased in the CGT group. The loricrin levels of the CGT group was 24,602±1090/20,000,000 pixel which was increased by 283% (P<0.05) compared with that of the AE group (Fig 3).
3. Formation of lipid barrier
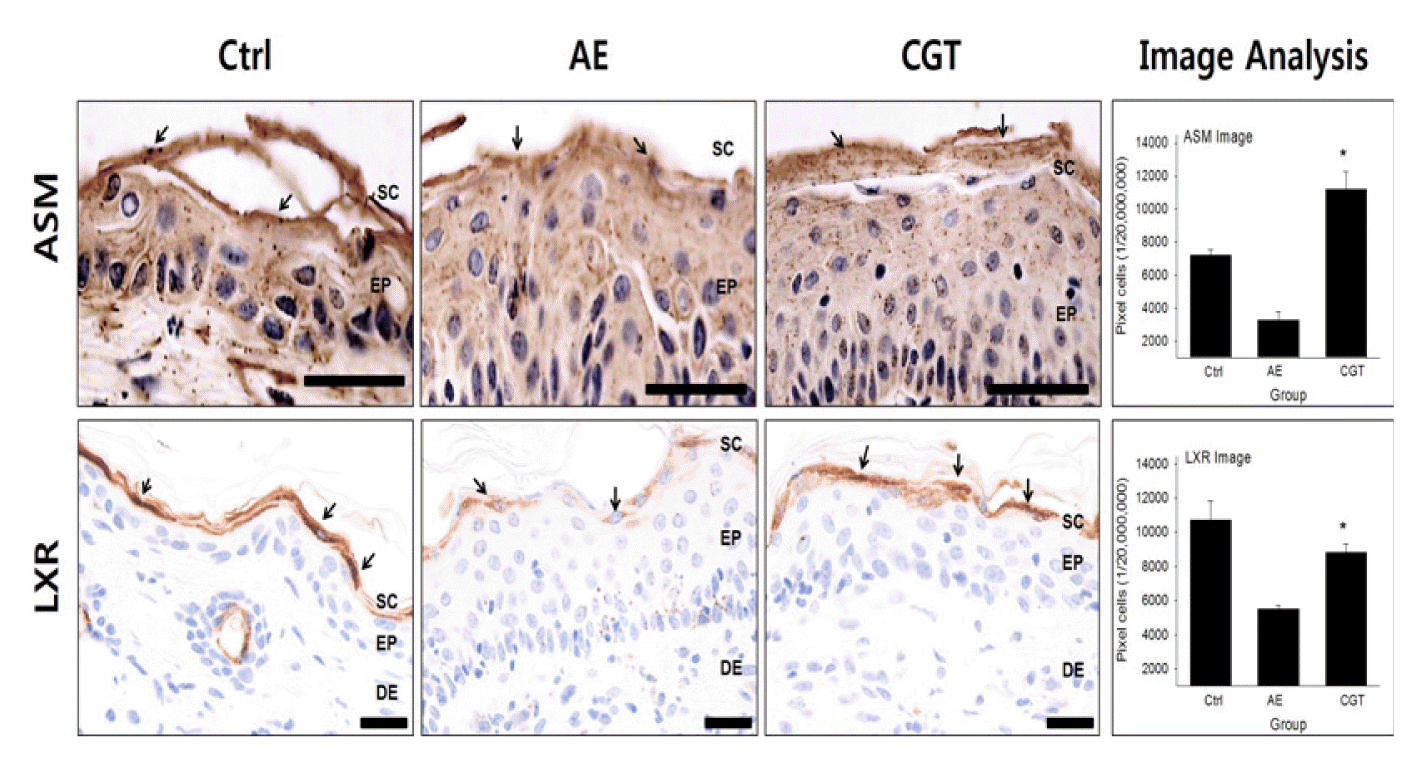
To investigate the generative effect of HTD treatment on the lipid barrier, ASM- and LXR-positive reactions were measured.
ASM-positive reaction in the stratum granulosum was 3,284±161/20,000,000 pixel which decreased by 93% in AE group compared with that of the Ctrl group. However, HTD treatments remarkably increased ASM levels in CGT group. The ASM level of the CGT group was 11,198±345/20,000,000 pixel which was increased by 240% (P<0.05) compared with that of the AE group (Fig 4).
LXR-positive reactions were widely exhibited in the cytoplasm of cells in the SC and stratum granulosum. Compared with the Ctrl group, the LXR level was 5,511±71/20,000,000 pixel which was remarkably decreased by 49% in the AE group. In contrast, both HTD treatments significantly improved LXR levels in the CGT group similarly to that of the Ctrl group. The LXR level of the CGT group was 8,840±149/20,000,000 pixel which was increased by 60% (P<0.05) compared with that of the AE group (Fig 4).
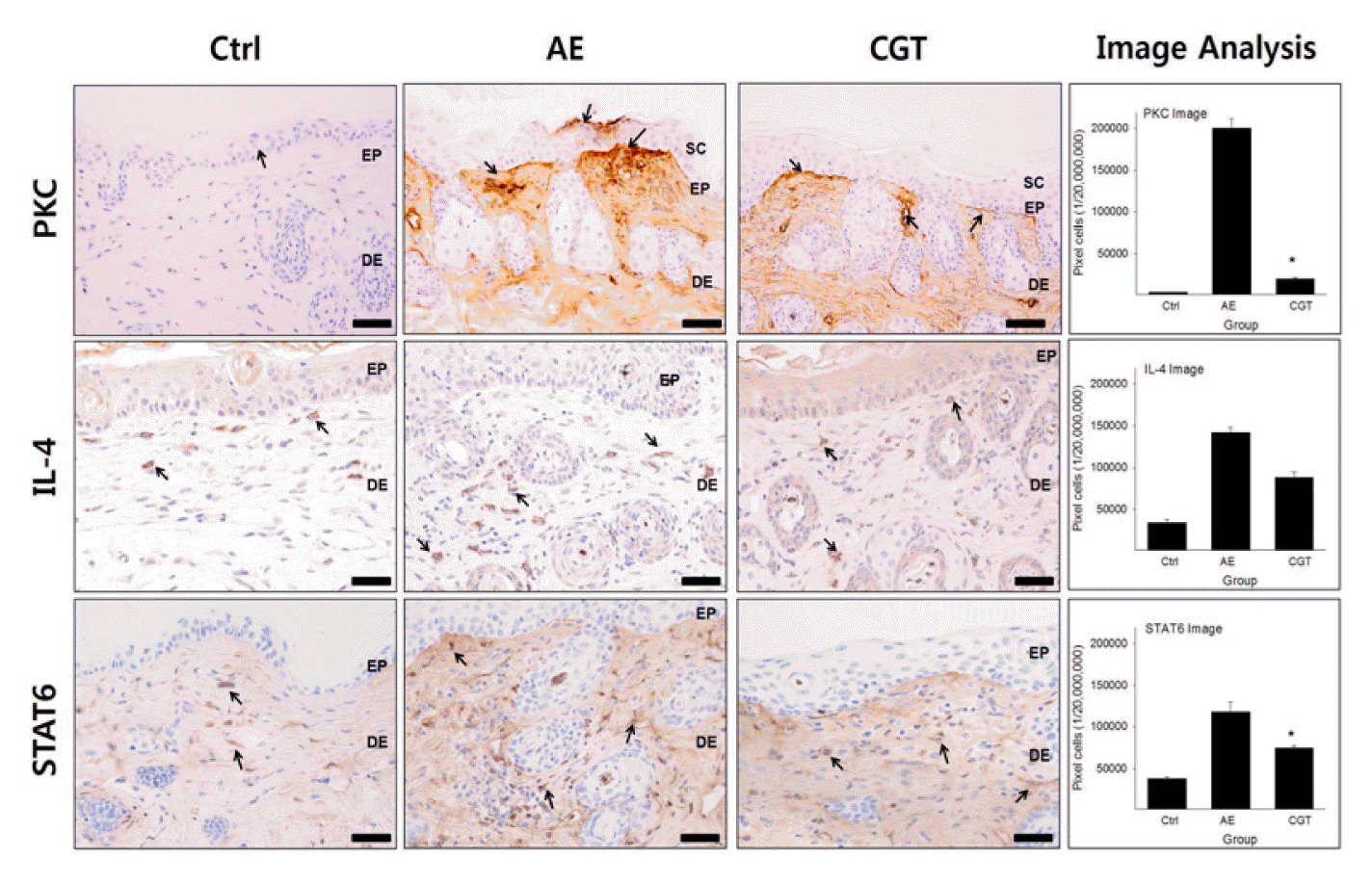
4. Regulatory effect on Th2 differentiation
PKC-, IL-4-, and STAT6-positive reactions were measured to observe the regulatory effect of HTD treatment on Th2 differentiation in the dermal papilla.
PKC immunohistochemistry was used to detect PKC-positive reactions in damaged epidermis and basement of epidermis. In the AE group, the level of PKC was 198,283±3590/20,000,000 pixel which was markedly increased by 5385% compared with that of the Ctrl group. However, HTD treatments also reduced the levels of PKC in the CGT group by 90% (P<0.05), compared with that of the AE group. The levels of PKC was 20,016±324/20,000,000 pixel in the CGT group (Fig 5).
The HTD treatments also remarkably reduced the levels of IL-4 and STAT6 compared with the AE group. The IL-4-positive reaction was 556,945±5852/20,000,000 pixel which was increased by 977% compared with that of the Ctrl group. In contrast, the levels of IL-4 in the CGT group was 114,731±4327/20,000,000 pixel which was decreased by 79% (P<0.05) compared with that of the AE group (Fig 5). Immunohistochemistry for STAT6 exhibited similar results as those for IL-4. In AE group, the STAT6-positive reaction was 117,672±3819/20,000,000 pixel which was increased by 210% compared with that of the Ctrl group. In contrast, levels of STAT6 in the CGT groups was 74,344±817/20,000,000 pixel which was decreased by 37% (P<0.05), compared with that of the AE group (Fig 5).
Discussion
In this study, we investigated the preventive effect of HTD treatment on AD-like skin lesions using DfE - induced obesity NC/Nga mice. H&E staining was observed to confirm the AD symptom relief effect of HTD treatments. HTD treatments was observed to alleviate capillary angiogenesis, edema and epithelial hyperplasia. We anticipated that relieved AD symptoms was due to well-maintained lipid barrier. To observe the effect of HTD treatments on maintaining the lipid barrier, changes of Lass2, filaggrin, involucrin, loricrin, ASM, and LXR levels were measured. HTD treatments helped to maintain the lipid barrier by increasing Lass2, filaggrin, involucrin, loricrin, ASM, and LXR levels. Moreover, we anticipated that HTD treatments relieved AD symptoms by contributing to the regulation of the Th2 predominant immune response, a major pathological mechanism of AD. To regulate Th2-skewed condition, HTD treatments affected Th2 differentiation by reducing PKC, IL-4, and STAT6 levels.
The hypothesis for etiopathogenesis of AD has shifted from “inside to outside” to “outside to inside”12). The “inside to outside” hypothesis suggests that the epidermal barrier is secondarily damaged due to rising inflammation caused by the imbalance between Th1 and Th2 cytokines, which is known as “Th2-skewed condition”13). However, the recent “outside to inside” hypothesis focuses on filaggrin. Filaggrin plays an important role in maintaining the SC by modulating hydration and pH level 14). The “outside to inside” hypothesis proposes that the defective SC due to impaired filaggrin stimulates the Th2 response by promoting persistent secretion of pro-inflammatory cytokines13, 14).
As mentioned earlier, obesity and inflammatory response are correlated. In particular, adipocytes produce and secrete cytokines such as TNF-α, IL-6, and monocyte chemotactic activating factor (MCP) -1, which play a central role in the inflammatory response15). Actually in obese subjects, an increase in pro-inflammatory cytokines such as TNF-alpha and IL-6 in the blood and an increase in CRP (c-reactive protein) as a biomarker of inflammatory diseases are observed16). Furthermore, many macrophages accumulate in adipocytes, leading to inflammation of the obese tissue itself, leading to chronic inflammation17).
Thus, we tried to confirm the efficacy of treatment of HTD treatments on AD by observing the levels of protein constituting lipid barrier in the obese conditions promoting the inflammatory response.
1. Alleviating effect on symptoms of AD
AD is characterized by pruritic skin lesions, impaired epidermal lipid barrier function, imbalance of the immune system, and allergic reactions18). Macroscopically, the HTD treatments alleviated AD symptoms such as edema including collagen fiber distribution and epithelial hyperplasia in obese NC/Nga mice. Furthermore, HTD treatment reduced angiogenesis. Angiogenesis of capillaries promoted by VEGFs is a hallmark of chronic inflammatory diseases such as AD. Angiogenesis is closely related to inflammation response by molecular links that cells involved in the inflammatory process release factors acting on the vascular endothelial cells19). Moreover, angiogenesis maintains inflammation by supplying oxygen and nutrients to cells in the inflammatory area20)
2. Maintenance of lipid barrier
Lass2, filaggrin, involucrin, and loricrin are closely related to maintenance of the epidermal lipid barrier. Ceramide is a typical substance to keep moisture and to form permeability barrier function in SC. Reduced ceramides in SC can cause skin abnormalities such as AD due to loss of protection against antigens, moisture loss from the epidermis, and dysfunction of barrier21). Among the Lass recently named ceramide synthase (CerS), Lass2 that catalyzes the synthesis of very long acyl chain ceramides is the most commonly expressed of all CerS and is most widely distributed in the human body22). Deficiency of ceramide synthesis containing heavy chain fatty acids by Lass2 causes severe skin diseases23). A reduced level of ceramide and changes in ceramide subclass were observed in AD or psoriasis24), and the replacement of ceramide to short chain lengths was reported in patients with AD25).
Filaggrin mutations causes skin barrier damage in AD and promotes IgE sensitization due to damaged skin barrier26). Although there are many causes for filaggrin reduction, particularly Th2-skewed condition contributes to reducing filaggrin levels in AD patient14). AD symptoms was relieved by improving the skin barrier due to increased expression of filaggrin27). Involucrin and loricrin are down-regulated by IL-4 in keratinocytes that process plays an important role in AD pathogenesis28). Particularly, decreased expression of involucrin and loricrin is inhibited in knock-out STAT6 mice, suggesting that down-regulation by IL-4 is mediated through a STAT6-dependent mechanism29).
In our previous study, we suggested that HTD treatment significantly increased filaggrin, involucrin, loricrin, and ASM levels, which are critical to the differentiation of epithelial cells and epidermal barrier function30). Consistent with previous findings, HTD treatments used in this study increased the level of Lass2, filaggrin, involucrin, and loricrin. Based on this result, HTD treatments may help to fortify the epidermal lipid barrier.
3. Formation of lipid barrier
ASM and LXR are closely related to formation of the epidermal lipid barrier. ASM, an enzyme that converts phospholipids into ceramides, also plays an important role in the formation of ceramides for epidermal barrier function31). ASM activity has been reported to be significantly reduced in AD lesions32), and reduced ASM activity is thought to be responsible for the reduced ceramide level, which is specific found in AD33). In addition, deficiency of ASM in AD inhibits recovery after damage to the skin barrier32) and the reduction of ASM activity in AD leads to skin barrier disorders by decreasing ceramide, involucrin, loricrin, and filaggrin34). In skin, activation of LXR improves permeability barrier homeostasis by stimulating epidermal lipid synthesis35), and promotes recovery of permeability barrier damage36). In addition, LXR controls cutaneous inflammation by inhibiting secretion of cytokine such as TNF-α and IL-1a that mediate the cutaneous inflammatory response37).
In this study, we compared the levels of ASM and LXR in order to examine the generative effect of HTD treatment on epidermal lipid barrier. The result showed that HTD treatment affected production of the levels of ASM and LXR. Based on this result, HTD treatments may help to generate the epidermal lipid barrier.
4. Regulatory effect on Th2 differentiation
Infiltrated external allergens can exacerbate Th2-skewed condition by activating inflammatory cells, including Th2 cells. For Th2 differentiation during inflammation, STAT6 signaling is an important pathway, and IL-4 is upstream of STAT638). STAT6 is closely related to IL-4 and IL-13 signaling, regulates important gene expression to IL-4 function and Th2 differentiation39), 40). In a knock-out STAT6 mice model, Th2 response by IL-4 does not occur41), and showed a loss of Th2 cell accumulation, which is one of the main features of asthma42). IL-4 is one of the key cytokines for Th2 differentiation and subsequent inflammation since it affects immune cells such as T cell, B cell, and macrophage through the α subunit of IL-4 receptor (IL-4Rα)38, 43). CD4 cells of knock-out IL-4 mice were unable to produce Th2 cytokines44). In additions, PKC is absolutely essential to differentiation of naive T cells into Th2 cells45) and PKC regulates the IL-4/STAT6 signaling pathway in T cells by phosphorylating Janus kinase (Jak) 146). PKC plays an important role in the Jak1/STAT6 signaling cascade associated with activation through IL-4 and Th2 differentiation. Moreover, PKC activation is important for epidermal inflammation since keratinocytes secreting inflammatory mediators such as TNF-α and cyclooxygenase-2 (COX-2) require PKC activation for appropriate differentiation47).
Based on these results, we compared the levels of IL-4, STAT-6 and PKC in order to examine the regulatory effect of HTD treatment on Th2 differentiation. In this study, HTD treatment affected reduction of the levels of IL-4, STAT6, and PKC. Down-regulation of IL-4 and PKC were also observed in our previous study48), but STAT6-inhibitory activity was reported here for the first time. Considering our results, HTD treatment may be effective for preventing inflammation induced by Th2-skewed condition by suppressing the main pathway of Th2 differentiation.
In previous studies, we demonstrated that HTD treatments reduce the inflammatory response by down-regulating Th2 differentiation48). In addition, we demonstrated that HTD treatments maintains and fortifies the lipid barrier11). Based on these results, we hypothesize that HTD could attenuate AD symptoms by maintaining lipid barrier in obese conditions, an aggravating factor of inflammation. In conclusion, the HTD treatments contributed to down-regulation of Th2 differentiation in AD-induced obese NC/Nga mice, leading to the alleviation of AD symptoms by maintaining the lipid barrier. Considering that the prevalence of childhood obesity is increasing, these results suggest that HTD treatments may contribute as an alternative treatment for the prevention of inflammatory diseases.