Introduction
Cerebral ischemia results from a variety of causes that cerebral blood flow is reduced due to a transient or permanent occlusion of cerebral arteries1). It is well known that transient global cerebral ischemia produces learning and memory impairment. Cerebral ischemia leads to neuronal cell death in the brain regions, such as the hippocampus, thalamus, cortex. and striatum2,3). Particularly, neurons in the hippocampal CA1 region are the most vulnerable cells to ischemic-reperfusion injury,4). Reduced neurons in the hippocampal CA1 region are associated with the defect in the memory function5).
Apoptosis plays a crucial role in the development and maintenance of homeostasis that may occur in multicellular organisms6). Inappropriate or excessive apoptosis has been implicated in the several types of neurodegenerative disorders, including ischemia7). Apoptosis is the most important process in CA1 neurons exposed to transient global ischemia8).
Cerebral ischemia is accompanied by reactive astrocyes and microglial activation in the hippocampal CA1 region9–11). Astrocytes, the most abundant glial cells in the brain, are known to associate closely with neuronal synapses12,13). However, reactive gliosis produces excess amounts of cytokines as well as inflammatory products that exacerbate ischemic damage14,15). Changes of glial cells may contribute to memory impairment16).
Cerebral ischemia increases activation of astrocyte and microglia in the hippocampus17). Glial fibrillary acidic protein (GFAP) is a major component of neurofilaments. Its overexpression is closely related to the morphological alterations of astrocytes in response to neuronal damage. Astrocyte activation is characterized by an increase in GFAP expression18). Li et al.19) reported that up-regulation of GFAP expression in a transient gerbil ischemia is a hallmark of ischemic brain injury.
Traditionally, acupuncture has been clinically applied for the treatment of various diseases and acupuncture possesses many effects, such as promotion of homeostasis, improvement in brain circulation, pain control, and neuromodulatory function20,21). Particularly, acupunctural treatment is effective for symptom improvement in cerebrovascular accidents, including ischemia15). Of these, acupuncture at ST40 acupoint traditionaly used to improve brain fuction (淸神志) and ameliorated the symptoms of Alzheimer’s disease22). Zhou and Jin22) reported that acupuncture at ST40 acupoint activated brain region which were closely correlated with the cognitive function, such as memory. However, the effect of acupuncture at the ST40 acupoint on apoptosis in relation with astrocytic reaction and microglial activation after transient global cerebral ischemia has not yet been reported. In the present study, we investigated the effect of acupuncture at ST40 acupoint on memory function and apoptosis in the hippocampal CA1 region following transient global cerebral ischemia in gerbils.
Materials and method
1. Experimental animals
Adult male Mongolian gerbils (12 – 14 weeks old) were used in this experiment. The experimental procedures performed with the Institutional Care and Use Committee (KHUASP[SE]-16-021) and conducted in accordance with the guiding principles for the care and use of animals approved by the Council of the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The gerbils were housed under controlled temperature (20 ± 2°C) and lighting (07:00 to 19:00 h) conditions with food and water provided ad libitum. Gerbils were randomly divided into six groups (n = 10 in each group): the sham-operation group(A), the sham-operation and ST40 acupoint-acupunctured group(B), the sham-operation and non-acupoint-acupunctured group(C), the ischemia-induction group(D), the ischemia-induction and ST40 acupoint-acupunctured group(E), the ischemia-induction and non-acupoint-acupunctured group(F). The same rules apply for the figures and tables shown below.
2. Induction of transient global ischemia
Transient global ischemia was induced with a previously described surgical procedure23). The gerbils were anesthetized with Zoletil 50® (10 mg/kg, i.p.; Vibac Laboratories, Carros, France). Following bilateral neck incisions, both common carotid arteries (CCA) were exposed and occluded with aneurysm clips for 5 min. The clips were then removed to restore cerebral blood flow. Body and rectal temperature were maintained at 36 ± 0.50°C during surgery using homeothermic blanket control unit (Harvard Apparatus, Massachusetts, MA, U.S.A.) that enveloped the body and the head. After recovery, animals were monitored for an additional 2 h to ensure that hypothermia did not occur. The animals in the sham-operation group were subjected to the same procedure, but the CCAs were not occluded following the neck incisions.
3. Acupuncture
The animals in the acupuncture treatment groups received acupuncture stimulation once daily (at 10:00 am) 7 consecutive days, starting one day after surgery. For acupunctural stimulation, stainless acupuncture needles of 0.3 mm diameter were inserted bilaterally into the ST40 acupoint that located at the midpoint lateral to the patella ligament from to the lateral malleolus of tibiofibula24), and into femoral region for the non-acupoint-treated group.
4. Step-down avoidance task
The latency of the step-down avoidance task was determined to evaluate short-term memory. Gerbils were trained in a step-down avoidance task 6 days after ischemia. One hour after training, the latency (s) of the animals in each group was determined. Gerbils were placed on a 7 × 25 cm platform 2.5 cm high. The platform faced a 42 × 25 cm grid of parallel 0.1 cm-caliber stainless steel bars spaced 1 cm apart. In training sessions, the animals received 0.5 mA, scramble foot shock for 2 s immediately upon stepping down. The interval of gerbils stepping down and placing all four paws on the grid was defined as the latency time. The latency over 180 s was counted as 180 s.
5. Tissue preparation
The gerbils were sacrificed immediately after determining the latency of the step-down avoidance task. The animals were anesthetized using Zoletil 50® (10 mg/kg, i.p.; Vibac Laboratories), transcardially perfused with 50 mM phosphate-buffered saline (PBS), and fixed with a freshly prepared solution consisting of 4% paraformaldehyde in 100 mM phosphate buffer (PB, pH 7.4). Brains were dissected, post-fixed in the same fixative overnight, and transferred to 30% sucrose for cryoprotection. Serial 40-μm-thick coronal sections were made cut with a freezing microtome (Leica, Nussloch, Germany).
6. Immunohistochemistry
Immunohistochemistry for GFAP-positive and CD11b-positive cells and for CD200-positive and CD200R-positive fibers in the hippocampus was performed. Free-floating tissue sections were first incubated in 3% H2O2 for 30 min to block the endogenous peroxidase activity. The sections were incubated in blocking solution (1% BSA and 10% horse serum in 0.05 M PBS) for 2 h at room temperature, and were then incubated overnight with anti-mouse GFAP antibody (1:1,000; Cell Signaling Technology, Beverly, MA, USA), anti-mouse CD11b antibody (1:500; Serotec, Raleigh, NC, USA), anti-goat CD200 antibody (1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), and anti-goat CD200R antibody (1:500; Santa Cruz Biotechnology) at 4°C. The next day, the sections were incubated for 1 h with biotinylated anti-mouse IgG (1:200; Vector Laboratories, Burlingame, CA, USA) and at room temperature, and were subsequently incubated with a Vector Elite ABC kit® (Vector Laboratories) for 1 h at room temperature. Finally, the tissues were washed in PBS, followed by a brief rinse in distilled water, and mounted individually onto slides. Slides were allowed to air dry and were then cover-slipped.
7. Immunofluorescence
Immunofluorescence for the NeuN-staining and caspase-3-staining cells in the hippocampus was performed. The sections were incubated overnight with anti-mouse NeuN antibody (1:500; Chemicon, Temecula, CA, USA) and anti-mouse caspase-3 antibody (1:400; Santa Cruz Biotechnology). The sections were next incubated for 2 h with fluorescein isothiocyanate (FITC)-conjugated anti-mouse secondary antibody (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) for the NeuN and caspase-3 antibody (1:400; Jackson ImmunoResearch Laboratories). The sections were then mounted on gelatin-coated glass slides, and the coverslips were mounted using fluorescent mounting medium (DakoCytomation, Carpinteria, CA, USA). The slides of the fluorescent images were captured using a confocal laser scanning microscopy (LSM-700; Carl Zeiss, München-Hallbergmoos, Germany).
8. TUNEL staining
To visualize DNA fragmentation, a marker of apoptosis, TUNEL staining was performed using an In Situ Cell Death Detection Kit® with Fluorescein (Roche, Mannheim, Germany). Sections were post-fixed in ethanol-acetic acid (2:1) and rinsed. Sections were then incubated with proteinase K (100 μg/ml), rinsed, and incubated in 3% H2O2, permeabilized with 0.5% Triton X-100, rinsed again, and incubated in the TUNEL reaction mixture. The sections were rinsed and visualized using label fluorescein-dUTP to the 39-end of DNA fragments. The coverslips were mounted using fluorescent mounting medium (DakoCytomation, Carpinteria, CA, USA). The slides of the fluorescent images were captured using a confocal laser scanning microscopy (LSM-700; Carl Zeiss).
9. Western blot analysis
Western blotting was performed to detect the expression of Bax and Bcl-2. Tissue samples harvested from the hippocampus were lysed in the protein lysis buffer containing 50 mM Tris-HCI (pH 7.5), 150mM NaCl, 0.5% deoxycholic acid, 1% nonidet-P40 (NP40), 0.1% sodium dodecyl sulfate (SDS), 1 mM phenylmethylsulfonyl fluoride (PMSF), and 100 μm/ml leupeptin. Protein concentration was measured using a colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein of 40 ug was separated on SDS-polyacrylamide gels and transferred onto a nitrocellulose membrane (Schleicher & Schuell GmbH, Dassel, Germany). Anti-mouse actin antibody (1:2000; Santa Cruz Biotechnology), anti-mouse Bax antibody (1:1000 Santa Cruz Biotechnology), and anti-mouse Bcl-2 antibody (1:1000 Santa Cruz Biotechnology) used as the primary antibodies. Horseradish peroxidase-conjugated anti-mouse antibody (1:2000; Santa Cruz Biotechnology) for actin, Bax and Bcl-2 were used as the secondary antibodies. Band detection was performed using as enhanced chemiluminescence (ECL) detection system (Amersham Pharmacia Biotech GmbH, Freiburg, Germany). To compare the relative expressions of proteins, the detected bands were calculated densitometrically using Image-Pro® Plus software (Media Cybernetics Inc., Silver Spring, MD, USA).
10. Data analysis
The numbers of NeuN-positive, GFAP-positive, and CD11b-positive positive cells in the hippocampal CA1 region (320 × 100 μm) were counted. Also, the optical densities of CD200-positive and CD200R-positive fibers were measured in 320 × 100 μm square images using an using Image-Pro® Plus software (Media Cybernetics Inc.). The results are expressed as mean ± standard error of the mean (S.E.M). SPSS Statistics version 20 (IBM SPSS, Chicago, IL, USA) was used for statistical analysis. Statistical analysis was performed using one-way ANOVA followed by Duncan's post-hoc test, and the results were expressed as the mean ± standard error of the mean (S.E.M). Significance was set as p < 0.05.
Results
1. Effect of acupuncture on the step-down avoidance task
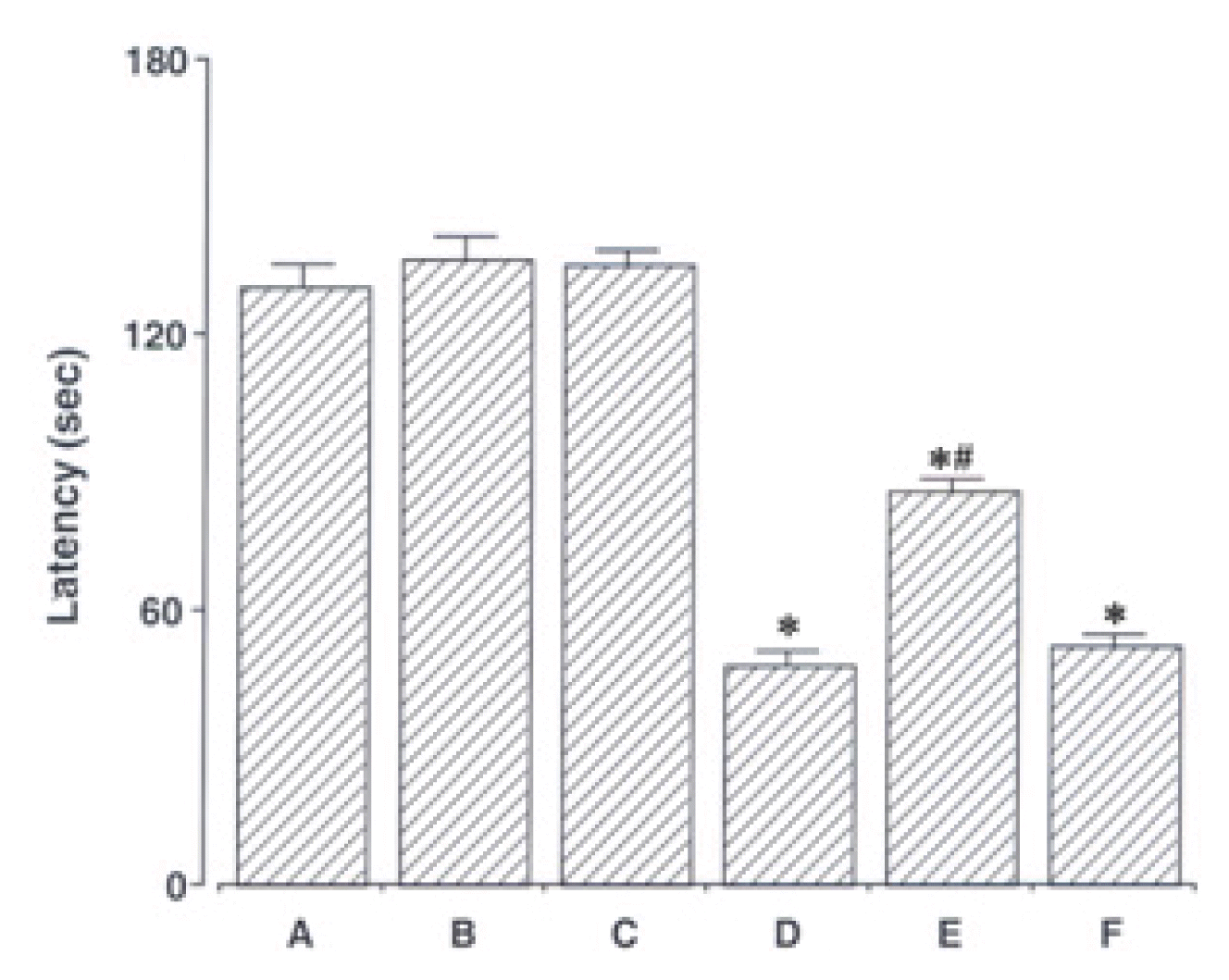
The latencies of the step-down avoidance task are presented in Fig. 1. The latency time (s) was 130.54 ± 4.73 sec in the sham-operation group, 136.72 ± 4.61 sec in the sham-operation and ST40 acupoint-acupunctured group, 135.36 ± 3.31 sec in the sham-operation and non-acupoint-acupunctured group, 47.90 ± 3.06 sec in the ischemia-induction group, 85.90 ± 2.38 sec in the ischemia-induction and ST40 acupoint-acupunctured group, 51.81 ± 2.34 sec in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that short-term memory was disturbed by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint alleviated ischemia-induced short-term memory impairment (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on short-term memory.
2. Effect of acupuncture on neuronal loss in the hippocampal CA1 region
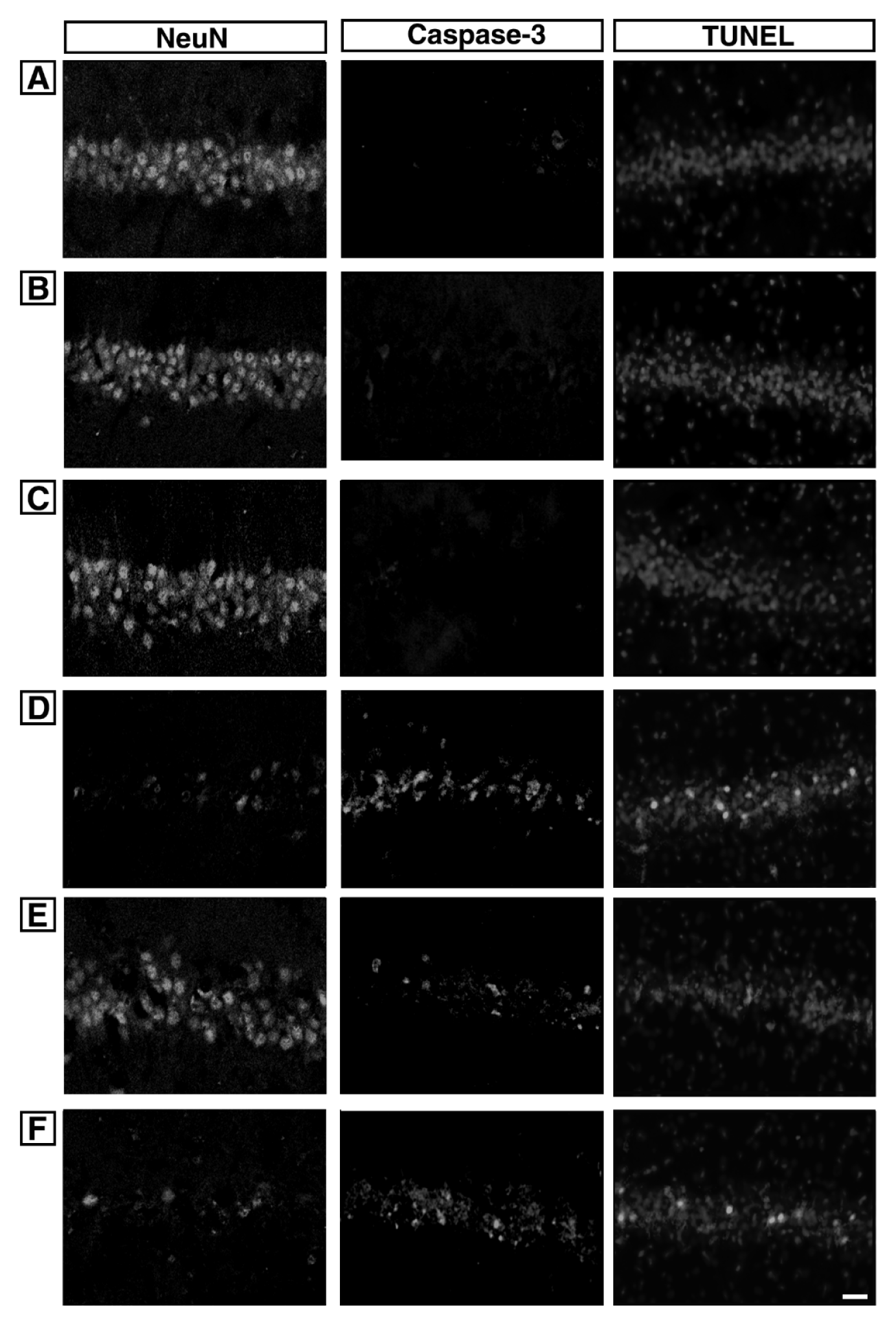
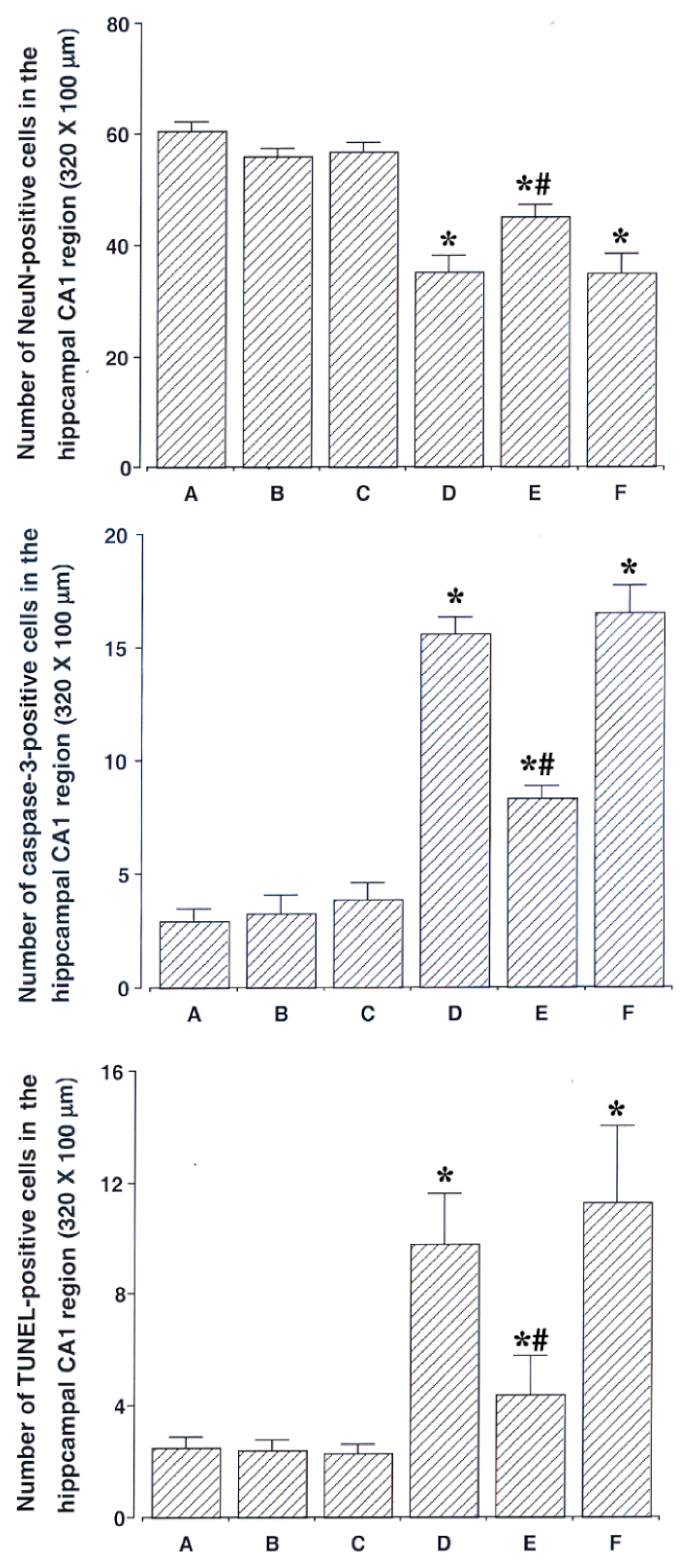
Photomicrographs of NeuN-positive cells in the hippocampal CA1 region are presented in Fig. 2. The number of NeuN-positive cells was 60.83 ± 1.42 in the sham-operation group, 56.17 ± 1.19 in the sham-operation and ST40 acupoint-acupunctured group, 56.94 ± 1.51 in the the sham-operation and non-acupoint-acupunctured group, 35.15 ± 2.81 in the ischemia-induction group, 45.00 ± 2.27 in the ischemia-induction and ST40 acupoint-acupunctured group, 35.00 ± 3.41 in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that the number of neurons in the hippocampal CA1 region was decreased by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint alleviated ischemia-induced neuronal loss in the hippocampal CA1 (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on neuronal loss.
3. Effect of acupuncture on caspase-3 and TUNEL-positive cells in the hippocampal CA1 region
Photomicrographs of caspase-3-positive cells and TUNEL-positive cells in the hippocampal CA1 region are presented in Fig. 2.
The number of caspase-3-positive cells was 2.90 ± 0.59 in the sham-operation group, 3.27 ± 0.81 in the sham-operation and ST40 acupoint-acupunctured group, 3.90 ± 0.68 in the sham-operation and non-acupoint-acupunctured group, 15.63 ± 0.69 in the ischemia-induction group, 8.36 ± 0.52 in the ischemia-induction and ST40 acupoint-acupunctured group, 16.54 ± 1.17 in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that caspase-3 expression in the hippocampal CA1 region was increased by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint alleviated ischemia-induced caspase-3 expression in the hippocampal CA1 region p (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on caspase-3 expression.
The number of TUNEL-positive cells was 2.50 ± 0.38 in the sham-operation group, 2.42 ± 0.43 in the sham-operation and ST40 acupoint-acupunctured group, 2.25 ± 0.25 in the sham-operation and non-acupoint-acupunctured group, 9.83 ± 1.83 in the ischemia-induction group, 4.40 ± 1.40 in the ischemia-induction and ST40 acupoint-acupunctured group, 11.33 ± 2.74 in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that DNA fragmentation in the hippocampal CA1 region was increased by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint alleviated ischemia-induced DNA fragmentation in the hippocampal CA1 region (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on DNA fragmentation.
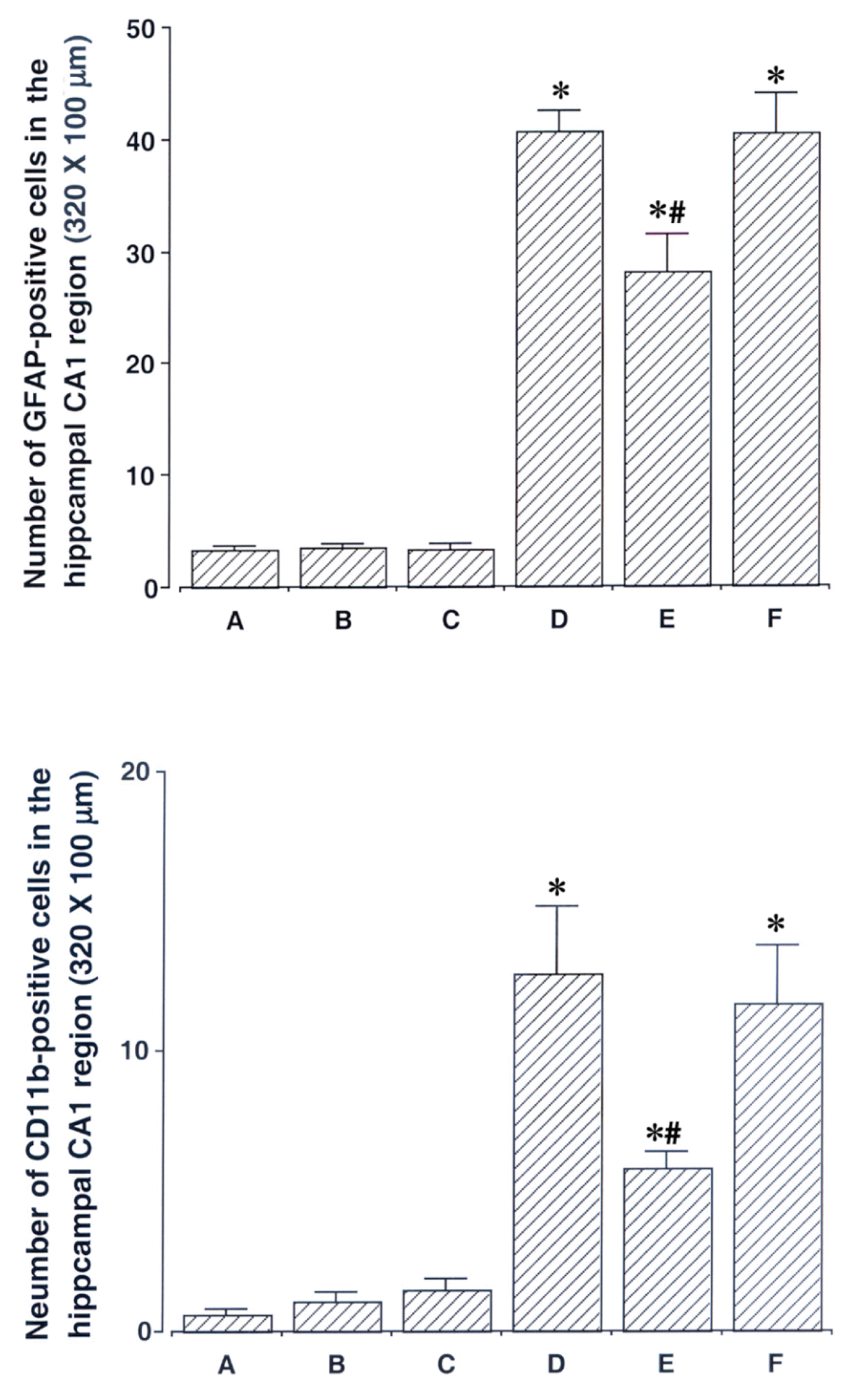
4. Effect of acupuncture on GFAP-positive cells in the hippocampal CA1 region
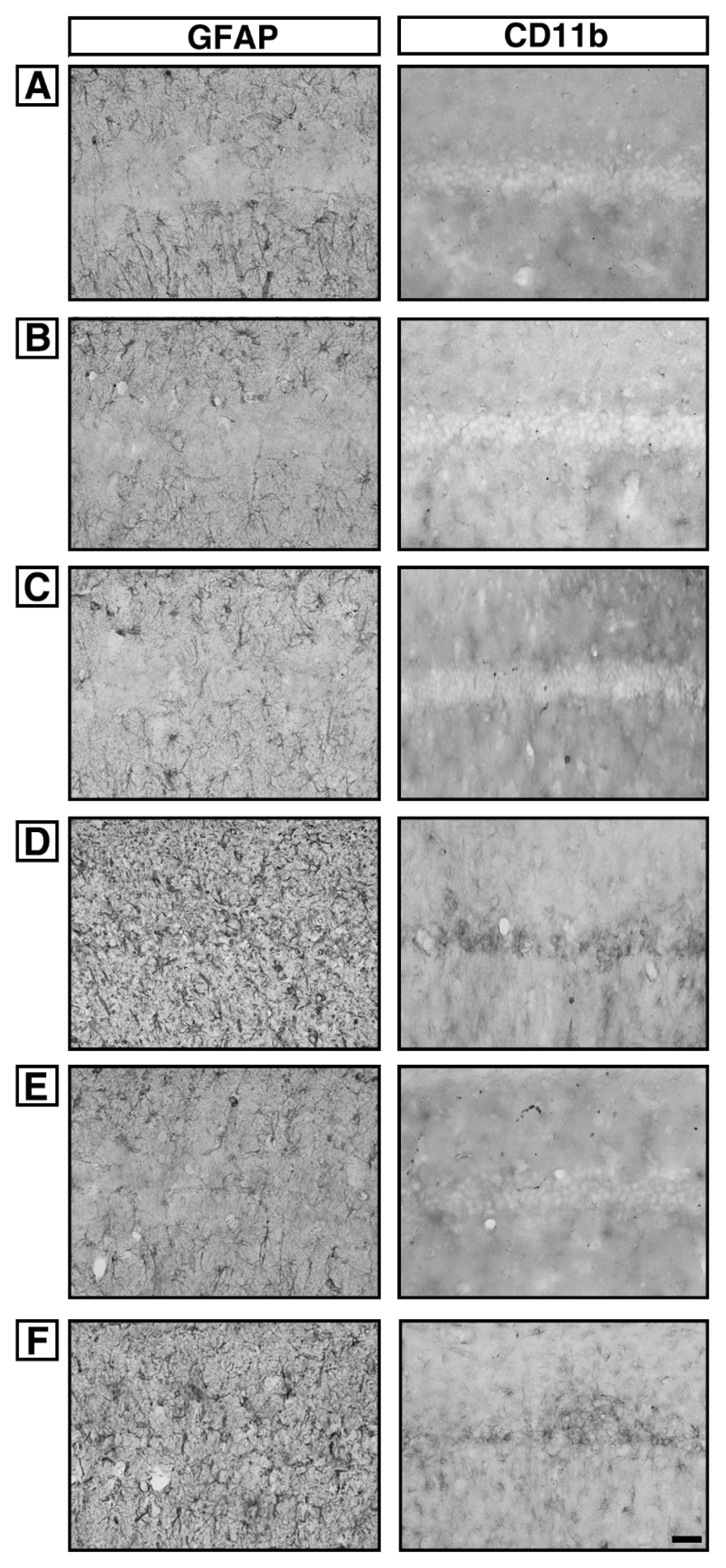
Photomicrographs of GFAP-positive cells in the hippocampal CA1 region are presented in Fig. 3. The number of GFAP-positive cells was 3.38 ± 0.31 in the sham-operation group, 3.53 ± 0.42 in the sham-operation and ST40 acupoint-acupunctured group, 3.41 ± 0.39 in the sham-operation and non-acupoint-acupunctured group, 40.64 ± 1.90 in the ischemia-induction group, 28.08 ± 3.43 in the ischemia-induction and ST40 acupoint-acupunctured group, 40.55 ± 3.43 in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that the number of GFAP-positive cells in the hippocampal CA1 region was increased by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint suppressed the number of GFAP-positive cells in the ischemic gerbils (p < 0.05). However, non-acupoint treatment did not exert significant effect on the number of GFAP-positive cells.
5. Effect of acupuncture on CD11b-positive cells in the hippocampal CA1 region
Photomicrographs of CD11b-positive cells in the hippocampal CA1 region is presented in Fig. 3. The number of CD11b-positive cells was 0.57 ± 0.17 in the sham-operation group, 1.13 ± 0.33 in the sham-operation and ST40 acupoint-acupunctured group, 1.50 ± 0.37 in the sham-operation and non-acupoint-acupunctured group, 12.77 ± 2.42 in the ischemia-induction group, 5.76 ± 0.57 in the ischemia-induction and ST40 acupoint-acupunctured group, 11.69 ± 2.12 in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that the number of CD11b-positive cells in the hippocampal CA1 region was increased by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint suppressed the number of CD11b-positive cells in the ischemic rats (p < 0.05). However, non-acupoint treatment did not exert significant effect on the number of CD11b-positive cells.
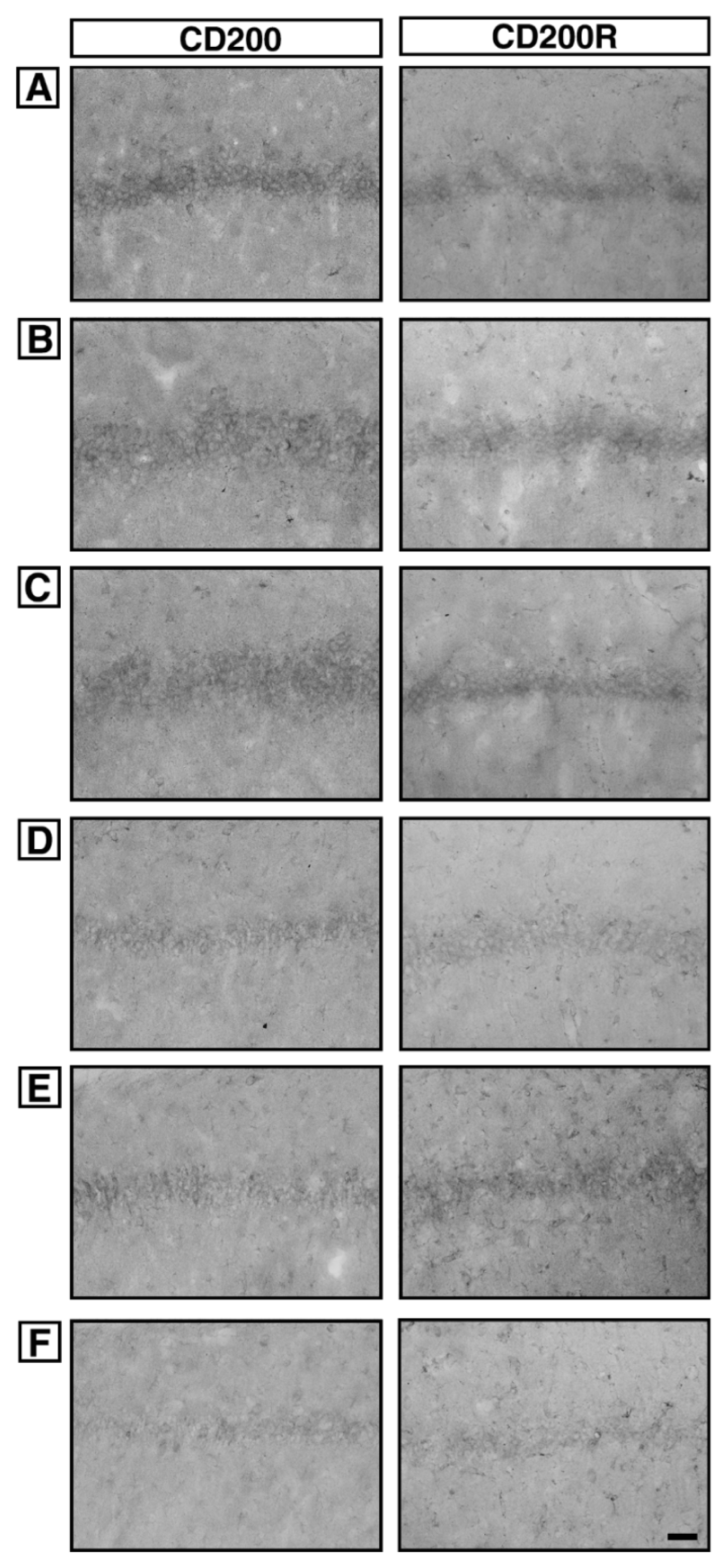
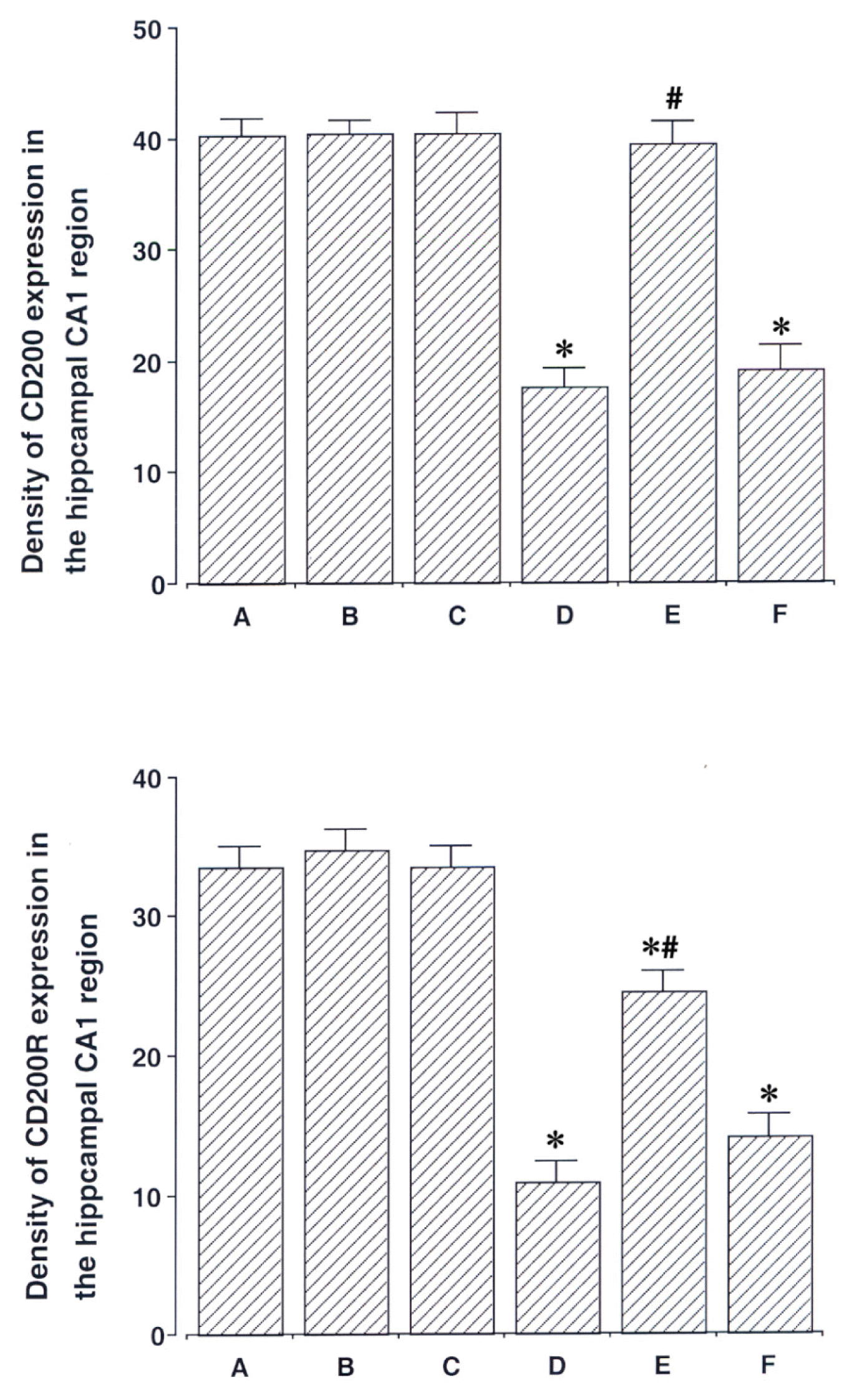
6. Effect of acupuncture on CD200 and CD200R expressions in the hippocampal CA1 region
Photomicrographs of CD200 and CD200R in the hippocampal CA1 region are presented in Fig. 4. The level of CD200 expression was 40.30 ± 1.53 in the sham-operation group, 40.50 ± 1.14 in the sham-operation and ST40 acupoint-acupunctured group, 40.45 ± 1.76 in the sham-operation and non-acupoint-acupunctured group, 17.64 ± 1.60 in the ischemia-induction group, 39.50 ± 1.95 in the ischemia-induction and ST40 acupoint-acupunctured group, 19.21 ± 2.07 in the ischemia-induction and non-acupoint-acupunctured group.
The level of CD200R expression was 33.49 ± 1.57 in the sham-operation group, 34.82 ± 1.43 in the sham-operation and ST40 acupoint-acupunctured group, 33.50 ± 1.50 in the sham-operation and non-acupoint-acupunctured group, 10.93 ± 1.38 in the ischemia-induction group, 24.51 ± 1.52 in the ischemia-induction and ST40 acupoint-acupunctured group, 14.12 ± 1.52 in the ischemia-induction and non-acupoint-acupunctured group.
These results showed that CD200 and CD200R expressions in the hippocampal CA1 region were decreased by induction of ischemic injury (p < 0.05). Acupuncture at ST40 acupoint increased CD200 and CD200R expressions in the ischemic gerbils (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on the CD200 and CD200R expressions.
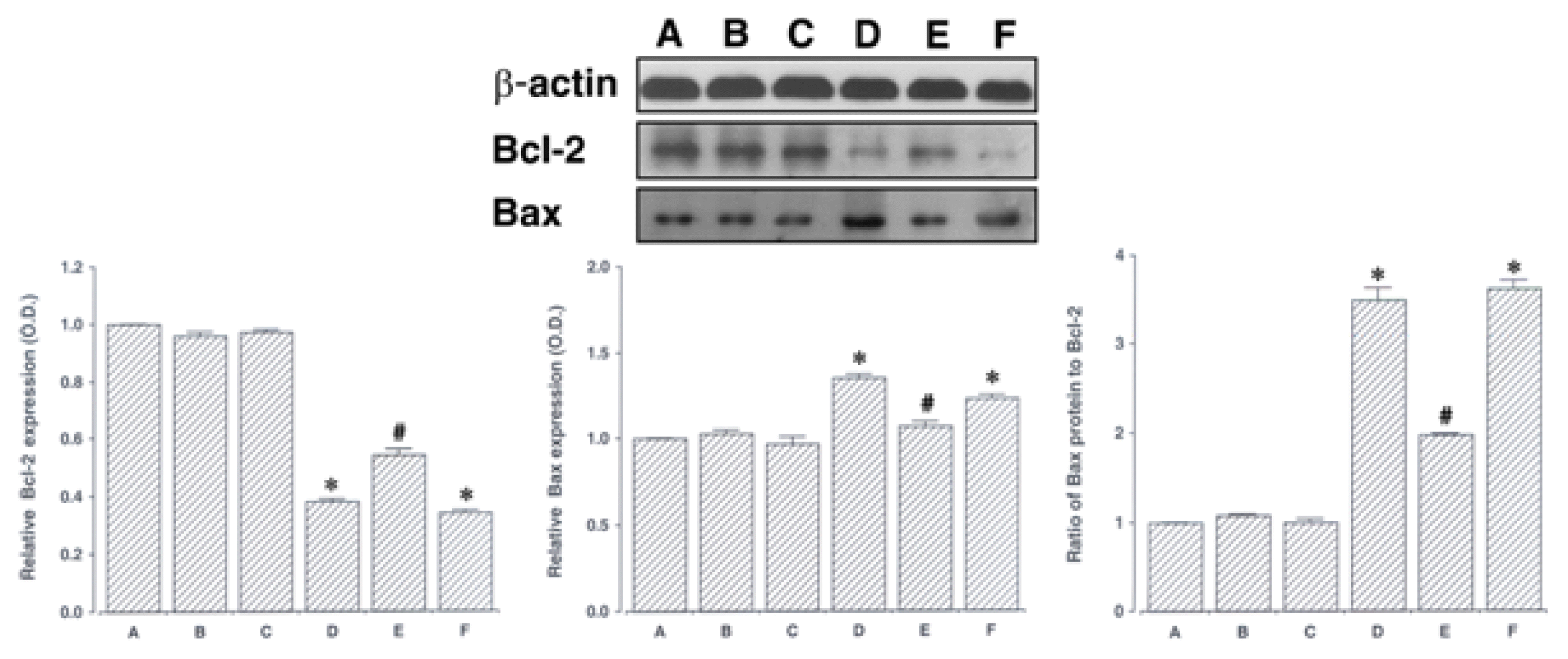
7. Effect of acupuncture on Bax and Bcl-2 expressions in the hippocampus
We determined the relative expression of Bcl-2 in the hippocampus (Fig. 5, lower left). When the level of Bcl-2 (26 kDa) in the sham-operation group was set at 1.00, the level of Bcl-2 was 0.96 ± 0.01 in the sham-operation and ST40 acupoint-acupunctured group, 0.97 ± 0.01 in the sham-operation and non-acupoint-acupunctured group, 0.38 ± 0.02 in the ischemia-induction group, 0.54 ± 0.02 in the ischemia-induction and ST40 acupoint-acupunctured group, and 0.34 ± 0.01 in the ischemia-induction and non-acupoint-acupunctured group.
The present results showed that the expression of Bcl-2 in the hippocampus was decreased by induction of ischemic injury (p < 0.05). In contrast, acupuncture at ST40 acupoint remarkably increased Bcl-2 expression in the ischemic gerbilss (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on the Bcl-2 expression.
We determined the relative expression of Bax in the hippocampus (Fig. 5, lower middle). When the level of Bax (21 kDa) in the sham-operation group was set at 1.00, the level of Bax was 1.03 ± 0.02 in the sham-operation and ST40 acupoint-acupunctured group, 0.97 ± 0.02 in the sham-operation and non-acupoint-acupunctured group, 1.35 ± 0.04 in the ischemia-induction group, 1.07 ± 0.15 in the ischemia-induction and ST40 acupoint-acupunctured group, and 1.23 ± 0.03 in the ischemia-induction and non-acupoint-acupunctured group.
The present results showed that the expression of Bax in the hippocampus was increased by induction of ischemic injury (p < 0.05). In contrast, acupuncture at ST40 acupoint remarkably suppressed Bax expression in the ischemic gerbils (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on the Bax expression.
The ratio of Bax to Bcl-2 was calculated (Fig. 5, lower right). When the ratio of Bax to Bcl-2 in the sham-operation group was set at 1.00, the ratio of Bax to Bcl-2 was 1.07 ± 0.02 in the sham-operation and ST40 acupoint-acupunctured group, 1.00 ± 0.03 in the sham-operation and non-acupoint-acupunctured group, and 3.49 ± 0.13 in the ischemia-induction group, 1.97 ± 0.03 in the ischemia-induction and ST40 acupoint-acupunctured group, and 3.61 ± 0.12 in the ischemia-induction and non-acupoint-acupunctured group.
The present results showed that the ratio of Bax to Bcl-2 in the hippocampus was enhanced by induction of ischemic injury (p < 0.05). In contrast, acupuncture at ST40 acupoint remarkably inhibited the ratio of Bax to Bcl-2 in the ischemic gerbils (p < 0.05). However, acupuncture at non-acupoint did not exert significant effect on the ratio of Bax to Bcl-2.
Discussion
Loss of hippocampal CA1 pyramidal neurons correlate with memory impairment in rats with ischemic insult25). Following transient global ischemia in gerbils, short-term memory in the step-down avoidance task was impaired23). In the present results, latency in the step-down avoidance task was shortened by induction of transient global ischemia, but this latency was improved by acupuncture at the ST40 acupoint. The present results indicate that acupuncture at ST40 acupoint alleviated short-term memory impairment induced by transient global ischemia in gerbils.
Two important groups of proteins involved in apoptotic cell death are the members of the Bcl-2 family and a class of cysteine proteases known as caspases. Bcl-2, an anti-apoptotic protein, is known to regulate apoptotic pathways and protects against cell death. Bax, a pro-apoptotic protein of that family, is expressed abundantly and selectively during apoptosis and promotes cell death. Increasing the ratio of Bax to Bcl-2 has commonly been used to determine the induction of apoptosis in several tissues26,27). Caspase-3 is one of the most widely studied members of the caspase families, and it is one of the key initiators of apoptosis28). Furthermore, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining is an assay that detects DNA fragmentation, which is the characteristic of apoptotic cell death. Commonly, neuronal apoptosis after global cerebral ischemia was observed by TUNEL staining29).
Deficit in learning and memory following cerebral ischemia has a close correlation with neuronal death in the hippocampal CA1 region 30). Hippocampal CA1 neurons are selectively vulnerable to global ischemia31). CA1 neuronal loss is estimated by NeuN staining, because neuN is a marker of mature neurons32). In the present study, neuronal loss in the hippocampal CA1 region was enhanced by induction of ischemia. In contrast, acupuncture at ST40 acupoint suppressed ischemia-induced neuronal loss in the hippocampal CA1 region.
Apoptosis is the major form of cell death in the brain after cerebral ischemia33). Apoptosis factors, such as TUNEL-positive cells and caspase-3-positive cells, in the brain were increased by transient global cerebral ischemia8,34). TUNEL staining shows DNF fragmentation representing apoptotic cell death after cerebral ischemia35). Caspase-3 is identified as a key mediator of apoptosis in animal models of ischemic stroke36). In the present results, the numbers of TUNEL-positive and caspase-3-positive cells in the hippocampal CA1 region were increased following ischemic insult, indicating that cerebral ischemia induced apoptotic cell death in the hippocampal CA1 region. In contrast, acupuncture at ST40 acupoint significantly decreased the numbers of TUNEL-positive and caspase-3-positive cell in the ischemic gerbils.
The Bcl-2 family proteins are distinct regulators for early stages of apoptosis37). Over-expression of Bax and down-regulation of Bcl-2 represent an active apoptotic process38). Over-expression of Bcl-2 and down-regulation of Bax represent anti-apoptotic process39). In the present results, Bax was increased and Bcl-2 was decreased in the hippocampal CA1 region following ischemic insult, indicating that cerebral ischemia initiated apoptotic cell death in the hippocampal CA1 region. In contrast, acupuncture at ST40 acupoint significantly decreased Bax expression and significantly increased Bcl-2 expression, thereby suppressed the ratio of Bax to Bcl-2 in the ischemic gerbils.
CD11b is the surface marker located on the plasma membrane of microglia and it is the β-integrin marker of microglia. CD11b expression represents microglial activation, and CD11b is increased during neurodegenerative inflammation40).
Reactive astrocytes are a hallmark of apoptotic cell death41). Apoptotic cell death in the brain is caused by activation of astrocyte and microglia42). Reactive astrocytes and microglial activation produces pro-inflammatory mediators and potentially neurotoxic compounds43). As a result, reactive astrocytes and microglial activation cause ischemic damage31,44). GFAP and CD11b represent reactive astrocytes and microglial activation, which are phenotype markers following brain ischemic injury45). In the present study, expressions of GFAP and CD11b in the hippocampal CA1 region were enhanced by induction of ischemia, representing reactive astrocytes and microglial activation initiated by ischemic insult. In contrast, acupuncture at ST40 acupoint suppressed expressions of GFAP and CD11b, which suggests inhibition of reactive astrocytes and microglial activation.
CD200 is a type 1 membrane glycoprotein and it is expressed on the surface of neurons. CD200 exerts its effect by binding to a CD200 receptor (CD200R) localized on microglia46). CD200 controls microglial activation47), and expressions of CD200 and CD200R were suppressed in the cerebral ischemia48).
Neuronal CD200 and glial CD200R signaling is implicated in microglial activation, which induces inflammation49,50). CD200-deficient mice exhibited microglial activation39). Loss of neuronal integrity causes breakdown of microglial inhibitory molecules, such as soluble inhibitor CD20051). In the present study, expressions of CD200 and CD200R in the hippocampal CA1 region were suppressed by induction of ischemia, representing activation of microglia. In contrast, acupuncture at ST40 acupoint enhanced expressions of CD200 and CD200R, which suggests inhibition of microglial activation.
Acupuncture showed a neuroprotective effect against cerebral ischemia in gerbils and monkeys16,52,53). One of the most impressive effects of the acupuncture is a rapid recovery from the complications of stroke54).
In the present study, acupuncture at ST40 acupoint ameliorated ischemia-induced short-term memory impairment by suppressing apoptosis in the hippocampus through down-regulation of reactive astrocytes and microglial activation. Thus the present study suggests that acupuncture at the ST40 acupoint can be used for treatment of patients with cerebral stroke.
Conclusion
We investigated the Effect of acupuncture on short-term memory and apoptosis after transient cerebral ischemia in gerbils.
Acupuncture at ST40 acupoint shows that
The latencies of the step-down avoidance task are increased.
Alleviated ischemia-induced neuronal loss in the hippocampal CA1.
Alleviated ischemia-induced caspase-3 expression in the hippocampal CA1 region.
Alleviated ischemia-induced DNA fragmentation in the hippocampal CA1 region.
Suppressed the number of GFAP-positive cells in the ischemic gerbils.
Suppressed the number of CD11b-positive cells in the ischemic rats.
Increased CD200 and CD200R expressions in the ischemic gerbils.
Increased Bcl-2 expression in the ischemic gerbilss.
Suppressed Bax expression in the ischemic gerbils.
Inhibited the ratio of Bax to Bcl-2 in the ischemic gerbils.
Acupuncture at ST40 acupoint ameliorated ischemia-induced short-term memory impairment by suppressing apoptosis in the hippocampus through down-regulation of reactive astrocytes and microglial activation. Thus the present study suggests that acupuncture at the ST40 acupoint can be used for treatment of patients with cerebral stroke.