Introduction
Phragmitis Rhizoma (PR) is dehydrated rhizoma of Phragmites communis Trinius (Gramineae). From ancient times, PR was used to treat fever and generate inner fluid. PR was recorded in Korean and Chinese pharmacopoeia and is still used by Oriental medicine physicians now. Useful effects of PR in pharmacopoeia are divided roughly into three. First, it alleviates fever of the lung and stomach. Second, it dissolves thirst due to high fever and generates inner fluid. Third, it stops vomiting and nausea. From the view of traditional Korean medicine, all of the above effects of PR are based on its heat-clearing activity. Heat-clearing activity in traditional Korea medicine represents many effects, including anti-inflammation effect. PR has traditionally been used to treat pulmonary tuberculosis, pulmonary edema, and pulmonary abscess by Oriental medicine physicians1).
However, there has been little scientific study of PR’s anti-inflammation effect before now. For the first step of scientific verification of PR’s anti-inflammation effect, this study was designed to measure secretion levels of inflammation cytokines (IL-6, IL-8, TNF-α), chemokines (RANTES, MCP-1) and histamines on the human alveolar epithelial cell line A549 (A549) and the human leukemic mast cell line (HMC) by CBA system and ELISA.
Materials and methods
1. Drugs
PR used in this study was purchased from Kwangmyungdang Medical Herbs, Korea. PR extract process was first, 300g of PR in 3,000ml DW heated in a heating extractor for 3 hours. Next, the extracts were filtered and concentrated using a rotary evaporator. Finally, the extracts were lyophilized using a freeze dryer (34.5g). The yield of extract was 11.5%.
2. Cell Culture
The human alveolar epithelial cell line (A549, Korean Cell Line Bank) was cultured in Roswell Park Memorial Institute medium 1640 (RPMI 1640) added in 10% bovine serum albumin, 2mM L-glutamine, 100IU/ml penicillin, 50ug/ml streptomycin at 37°C, 5% CO2, and 95% humidity. The human leukemic mast cell line (HMC) was cultured in Iscove’s Modified Dulbecco’s Media (IMDM) added in same the treatment and conditions as the A549. The cells were passaged every 2~3 days.
3. Treatment and Stimulation
A549 cells were in 24-well plates and stabilized for 24 hours, then divided into four groups; blank, control, high concentration reagent treated group (HG), and low concentration reagent treated group (LG). PR 300 (HG) and 30 (LG) ug/ml were treated on each plate well except in the blank group; phosphate buffered saline (PBS) was treated to the blank group wells. After 12 hours, Control, HG, and LG cells were stimulated by TNF-α (10ng/ml) and IL-4 (10ng/ml); the same volume of PBS was treated to the blank group wells. After 24 hours, the supernatant of each well medium was gathered and frozen at −20°C. The treatment of HMC was the same as all of the A549 conditions, except stimulation time and compound. Compound 48/80 (1 mg/ml) was treated for 15 minutes in HMC stimulation. The supernatants of HMC were gathered and directly used to measure histamine level.
4. Histamine Concentration Assay
Histamine level was measured by OPA-spectrofluorimetry. Specific processes of histamine assay were as follows. First, standards were made by serial dilution (top=1 mg/ml). Second, NaOH (1M) was added to the samples and standard wells. Third, o-phthaldialdehyde (1mg/ml) was added to all wells. After 4 minutes, fluorescence absorbance of samples and standards were checked by ELISA measuring instrument Synergy HT (Bio-Tek Instruments INC., USA).
5. Human Chemokine Assay
Human chemokine levels in culture supernatants of A549 were measured by a cytometry bead array (CBA). Interleukin-8 (CXCL8/IL-8), RANTES (CCL5/RANTES), monokine-induced by interferon-γ (CXCL9/MIG), monocyte chemoattractant protein-1 (CCL2/MCP-1), and interferon-γ-induced protein-10 (CXCL10/IP-10) levels were quantitatively measured using BD CBA Human Chemokine Kit I (BD biosciences, USA). In this study, only MCP-1, RANTES, and IL-8 were detected in A549 cells. All CBA procedures were according to manufacturer’s guide protocol. BD FACS Calibur (BD Bioscience, USA) was used in this experiment.
6. MTT assay for cell viability
General viability of cultured cells was determined by reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) to formazan. A549 cells were seeded onto a 24 well-plate and cultivated in DMEM medium for 24 hours, supplemented with samples (High conc. 300ug/ml, Low conc. 30ug/ml), and incubation was continued for an additional 24 hours. After addition of MTT solution, A549 cells were incubated for 4 hours at 37°C. The crystal of MTT was dissolved by DMSO, and the absorbance was measured at 595 nm by ELISA reader. The MTT method for HMC was as same as with A549, but incubation time was 8 hours.
Results
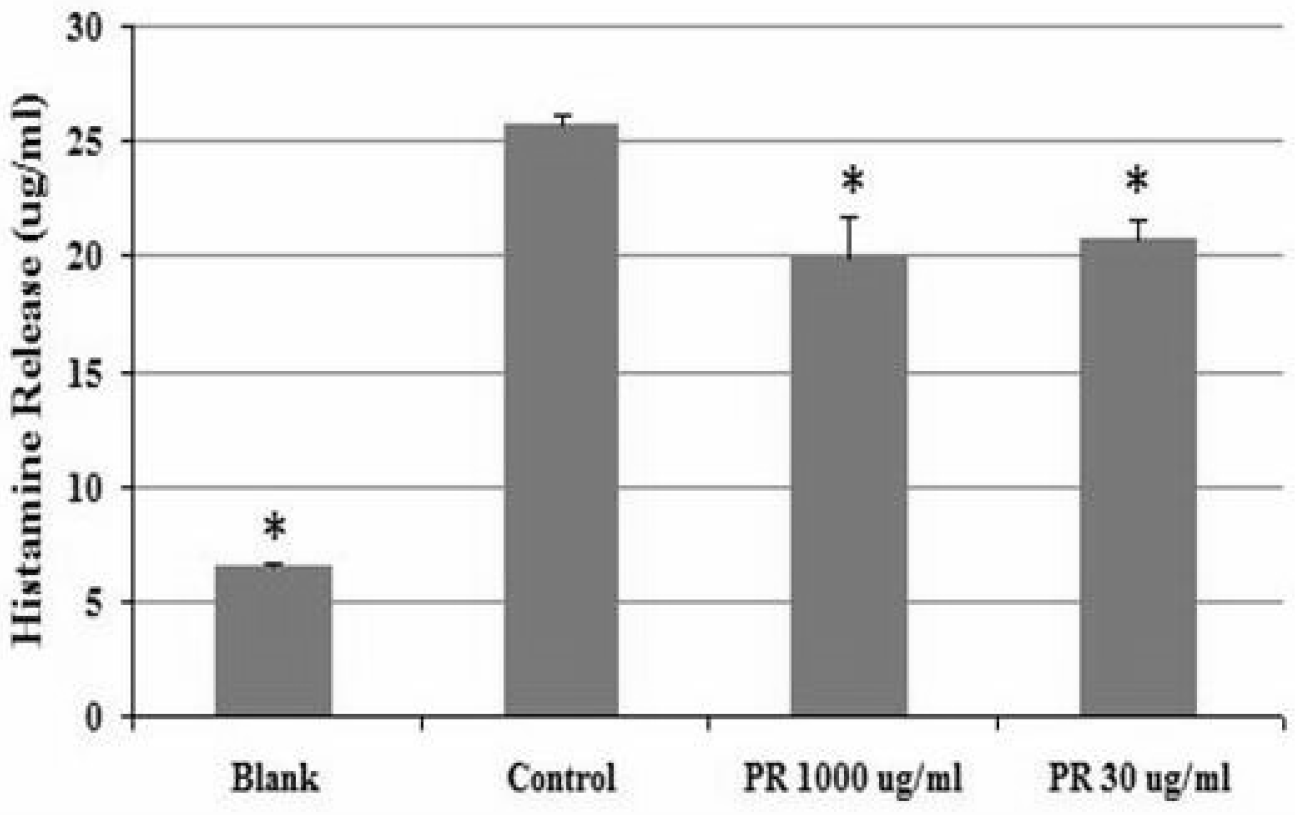
1. Effect of Phragmitis Rhizoma on histamine secretion
The histamine secretion of the blank group was 6.51±0.07pg/ml, and of the control group was 25.67 ±0.52pg/ml. The histamine secretion of the PR 1000 ug/ml treated groups was 19.88±1.79pg/ml, and of the PR 30ug/ml treated groups was 20.80±0.76 pg/ml (p<0.05). PR (1000ug/ml and 30ug/ml) showed a decrease of histamine releases compared to control (p<0.05, Figure 1).
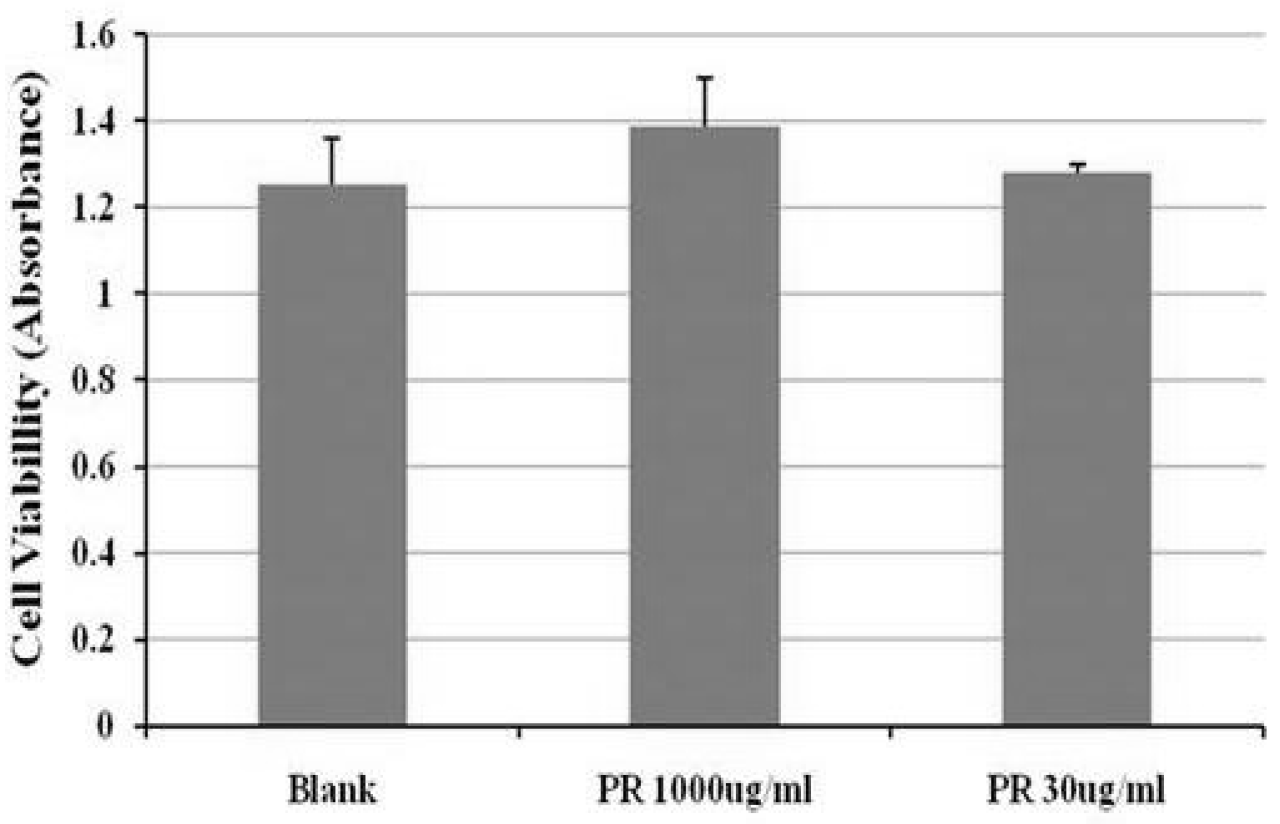
2. Cell viability of HMC
General viability of cultured HMC against PR was determined by reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) to formazan.
The cell viability of the blank group (12 h) was 1.25±0.11pg/ml. The cell viability of the PR 1000 ug/ml treated groups (12 h) was 1.39±0.11pg/ml, and of the PR 30ug/ml treated groups (12 h) was 1.28 ±0.02pg/ml (Figure 2.)
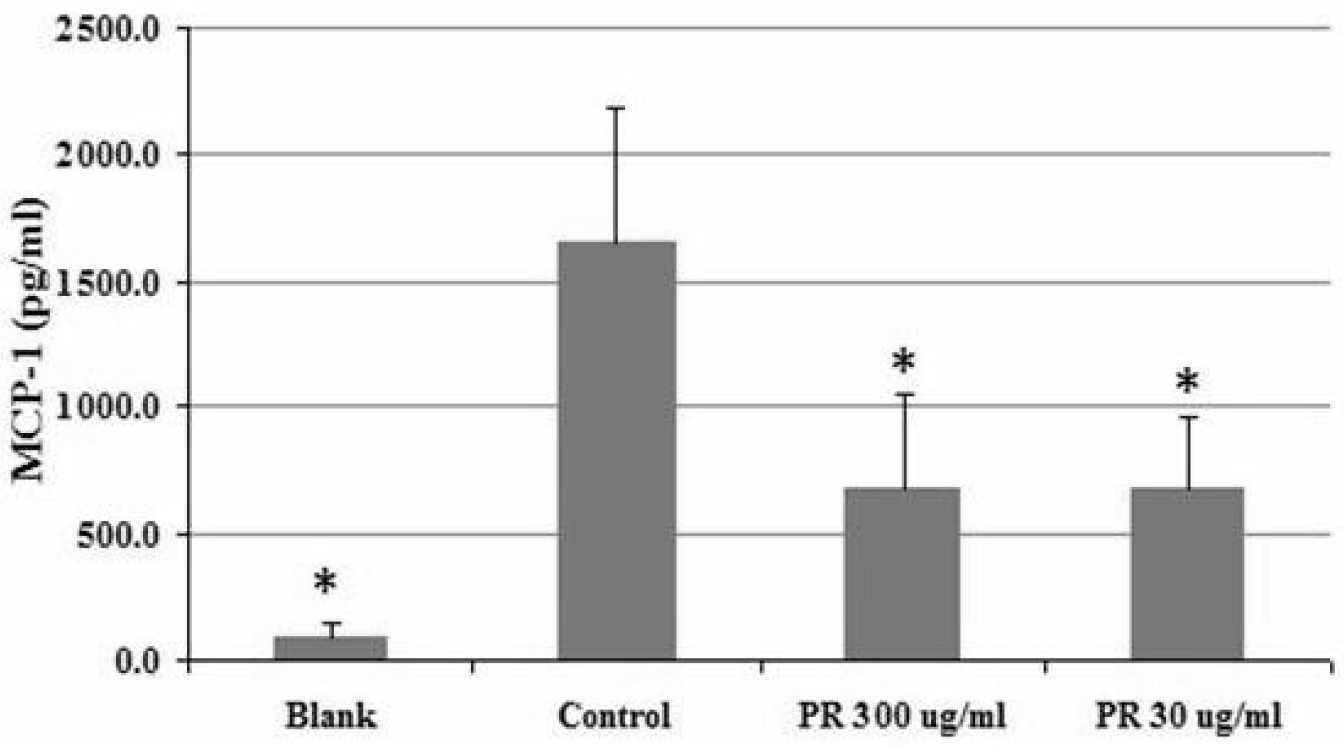
3. Effects of Phragmitis Rhizoma on the expression of chemokines
Inhibiting ratios of chemokines were measured using CBA software and each sample was calculated for inhibiting ratios.
The MCP-1 expression of the blank group (24h) was 92.4±49.7pg/ml, and of the control (24h) was 1650.4±531.13pg/ml. The IL-8 expression of the PR 300ug/ml treated groups (24h) was 675.6±372.45 pg/ml (p<0.05), and of the PR 30ug/ml treated groups (24h) was 676.0±279.66pg/ml (p<0.05, Figure 3). PR significantly decreased the MCP-1 expression in both the 300 and 30ug/ml treated conditions.
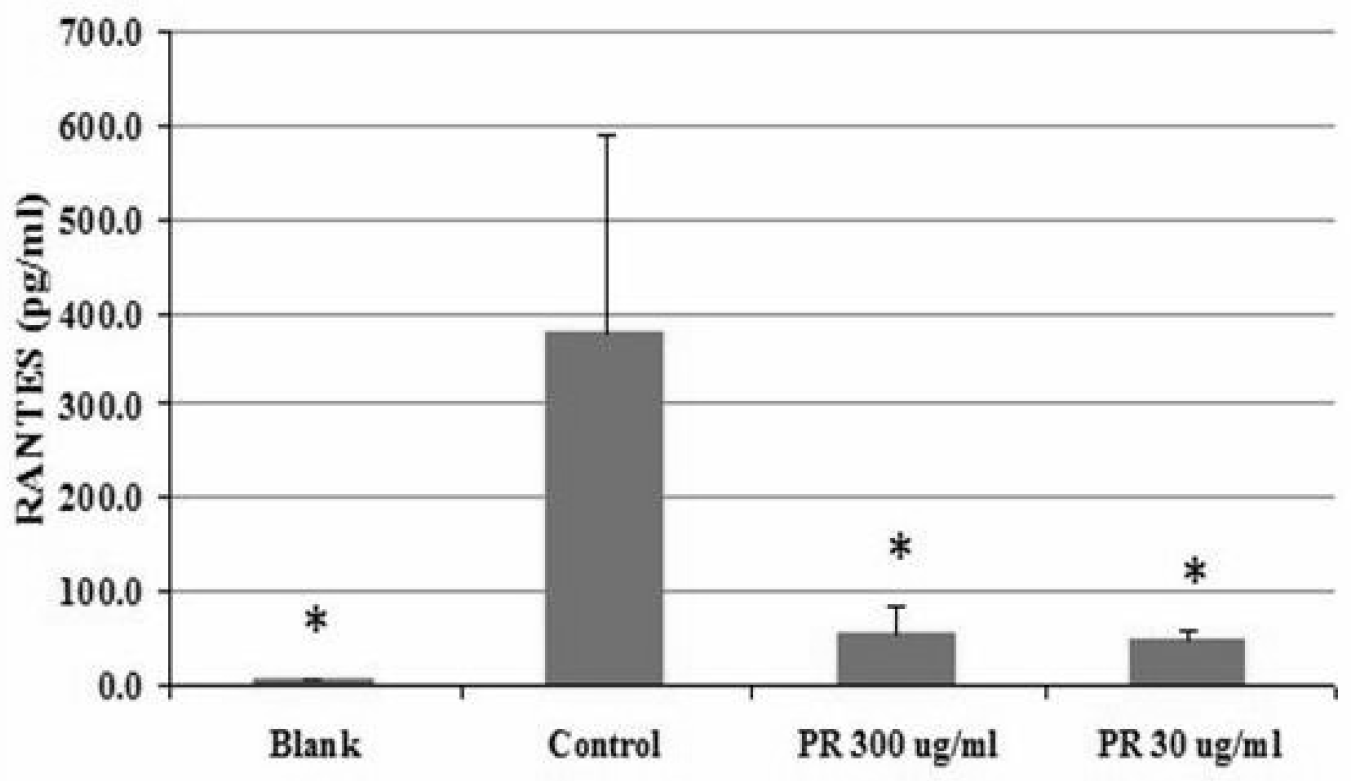
The RANTES expression of the blank group (24 h) was 6.4±0.77pg/ml, and of the control (24h) was 379.4±210.15pg/ml. The RANTES expression of the PR 300ug/ml treated groups (24h) was 53.6±32.22 pg/ml (p<0.05, Figure 4), and of the PR 30ug/ml treated groups (24h) was 48.1±9.42pg/ml (p<0.05, Figure 4). PR significantly decreased the RANTES expression in both the 300 and 30ug/ml treated conditions.
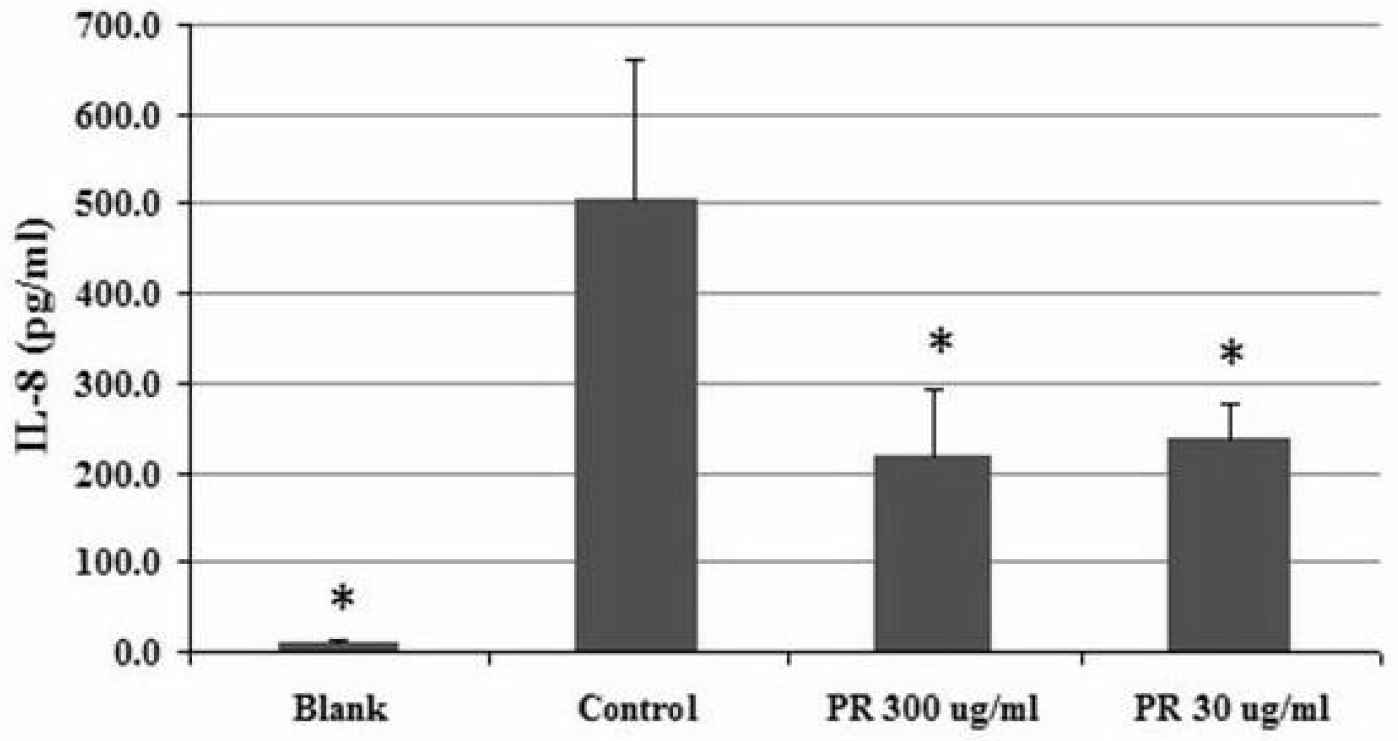
The IL-8 expression of the blank group (24h) was 9.4±2.16pg/ml, and of the control group (24h) was 503.8±157.58pg/ml. The IL-8 expression of the PR 300ug/ml treated groups (24h) was 219.6±73.30 pg/ml (p<0.05, Figure 5), of the and PR 30ug/ml treated groups (24h) was 238.8±39.30pg/ml (Figure 5). PR significantly decreased the IL-8 expression in both the 300 and 30ug/ml treated conditions.
4. Cell viability of A549 cells
General viability of cultured A549 cells against PR was determined by reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) to formazan.
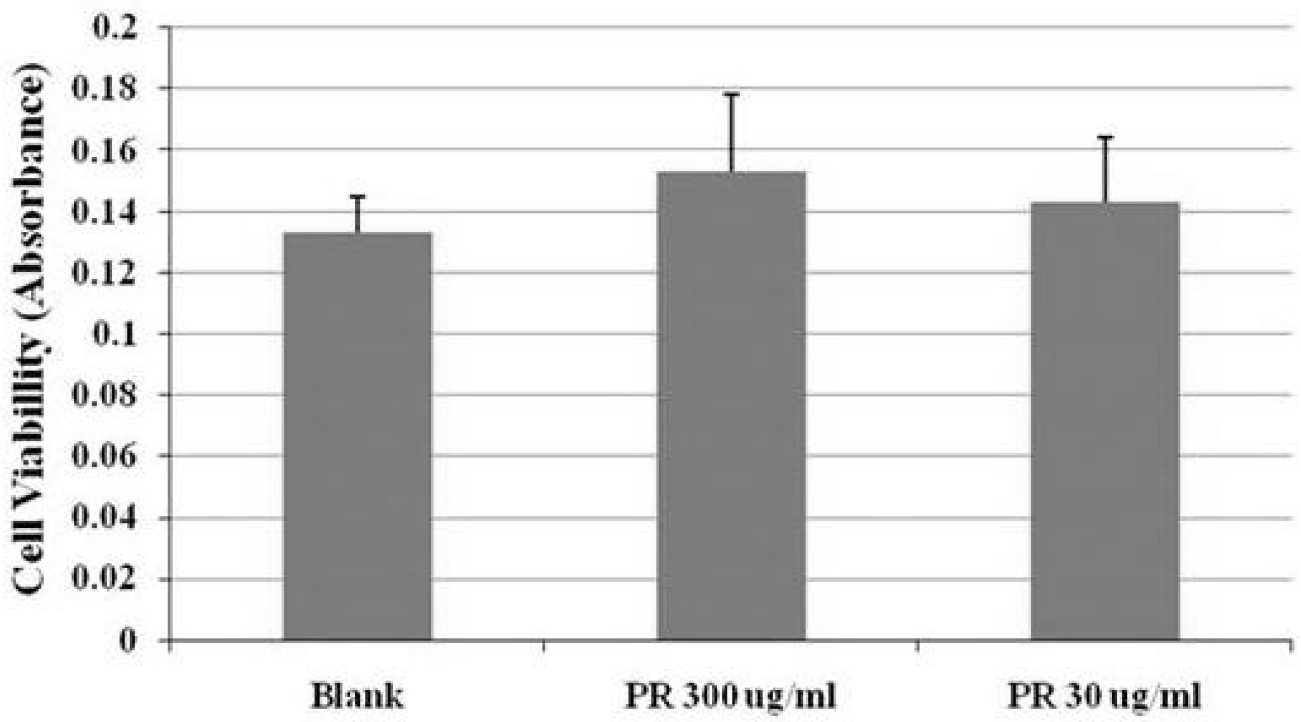
The cell viability of the blank group (28 h) was 0.13±0.01pg/ml. The cell viability of the PR 300 ug/ml treated groups (28 h) was 0.15±0.03pg/ml, and of the PR 30ug/ml treated groups (28 h) was 0.14±0.02pg/ml (Figure 6).
Discussion
Nowadays there are few studies about physiology activity of Phragmitis Rhizoma. However, several reports about hyperlipemia, diabetes and skin whitening have been made in Korea. Lee reported on the effect of PR on synthesis of glucose, insulin and lipids in blood serum2). Kim reported a suppressing action of increasing of glucagon granulation in Langerhans islet α cells with streptozotocin administration and suppressing effect of insulin degranulation in β cells3). Choi et al. reported MeOH extract of PR has lowering effects on cholesterol and glucose in the blood4). Choi et al. reported that MeOH extract of PR makes concentration of triglyceride about hypertriglyceridemic, hypercholesterolemic and diabetic hyperlipidemic rats, and β-sitosterol and ρ-coumaric acid parted from MeOH extract of PR have an effect that improves concentration of fat in the blood5). Jang mentioned PR’s effects of whitening and nutrition supply of skin6). Yoon reported that PR’s phenolic compounds have whitening activity on the B16 melanoma cell7).
Cytokines are small secreted proteins which mediate and regulate immunity, inflammation, and hematopoiesis. They must be produced de novo in response to an immune stimulus. They generally act over short distances and short time spans and at very low concentration. They act by binding to specific membrane receptors, which then signal the cell via second messengers, often tyrosine kinases, to alter its gene expression. Responses to cytokines include increasing or decreasing expression of membrane proteins, proliferation, and secretion of effector molecules.
It is common for different cell types to secrete the same cytokine or for a single cytokine to act on several different cell types. Cytokines are redundant in their activity, meaning similar functions can be stimulated by different cytokines. Cytokines are often produced in a cascade, as one cytokine stimulates its target cells to make additional cytokines. Cytokines can also act synergistically or antagonistically8,9,10).
IL-6 is an interleukin that acts as a pro-inflammatory. It is secreted by T cells and macrophages to stimulate immune responses to trauma, especially burns or other tissue damage leading to inflammation. IL-6 is one of the most important mediators of fever and of the acute phase response. IL-6 can be secreted by macrophages in response to specific microbial molecules, referred to as pathogen-associated molecular patterns (PAMPs). These PAMPs bind to highly important detection molecules of the innate immune system, called toll-like receptors (TLRs), that are present on the cell surface (or in intracellular compartments) which induce intracellular signaling cascades that give rise to inflammatory cytokine production11).
IL-8 is the most extensively studied member of the chemokine superfamily, with its major actions being as a neutrophil chemoattractant and activator. It has been reported to play a major role in triggering and sustaining the allergic inflammatory response12).
Monocyte chemotactic protein-1 (MCP-1) is a member of the C-C branch and is thought to play an important role in the allergic inflammatory process. The activities of MCP-1 are restricted to monocytes, lymphocytes13,14), and basophils15). MCP-1 has been reported to be increased in nasal washings of ragweed-sensitive subjects during the pollen allergen season16). Immunoreactivity for MCP-1 is also increased in the bronchial epithelium of asthmatic patients17).
Regulated upon Activation, Normal T-cell Expressed, and Secreted (RANTES or CCL5) is a protein classified as a member of the IL-8 superfamily of cytokines. It is known that production of RANTES could contribute to the inflammation that characterizes the respiratory tract in asthma and rhinitis18). Recently, increased levels of RANTES have been detected in the nasal secretions of atopic patients exposed to local allergen challenge19).
Histamine is a substance secreted from mast cells when an outward substance is detected by the immune system as a foreign substance. If secreted in quantity, an allergic reaction arises. The release of histamine causes several allergic symptoms. First, it distributes to an inflammatory response. Second, it causes constriction of smooth muscles. Histamine can cause inflammation directly as well as indirectly. Upon release of histamine by an antigen activated mast cell, permeability of vessels near the site is increased. Thus, blood fluids enter the area causing swelling. This is accomplished due to histamine’s ability to induce phosphorylation of an intercellular adhesion protein found on vascular endothelial cells. Indirectly, histamine contributes to inflammation by affecting the functions of other leukocytes in the area. It has been suggested by Marone et al. that histamine release triggers the release of cytokines and inflammatory mediator by some neighboring leukocytes. These chemicals in turn increase the inflammatory response20,21,22,23).
In the PR-treated group, the expression of interleukin 8 (IL-8), monocyte chemoattractant protein-1 (MCP-1), and chemokine (C-C motif) ligand 5 (Ccl5, RANTES) decreased significantly. These results support that treatment with PR can reduce the levels of IL-8, MCP-1, RANTES in A549 cells.